
Neuronšō╬─ĮŌūxŻ║┤¾╩¾īŹ“ד×ūC│╔░a┤µį┌ąįäe▓Ņ«É
“╚²╩Õ╩ŪéƤ¤²g│¼╦─╩«─ĻĄ─└Ž¤¤śī┴╦Ż¼ę╗ą”Š═┬Č│÷ā╔┼┼┤¾³Sč└Ż¼š¹╚š¤¤▓╗ļx╩ų”
“┤Õ└’└Ž└Ņ╩ŪéĆŠŲ╣ĒŻ¼├┐▓═Č╝ę¬║╚╔Žę╗┐┌Ż¼╗▄ć╔Ž│į┼▌├µČ╝▒žĒÜš¹Ų┐Č■ÕüŅ^”
“ÓÅŠė╝ęĄ─ąĪ║óĮą░óÅŖŻ¼į┌ūx│§ę╗Ż¼│┴─ńė┌ŠWĮjė╬擤oĘ©ūį░╬Ż¼ÓÅŠė┤¾ĮŃ×ķ┤╦ę╗╗I─¬š╣”
šfŲ│╔░aŻ¼─Ń─Xūė└’┐╔─▄Ģ■┴ó±R├░│÷└Ž¤¤śī╚²╩ÕĪóŠŲ╣Ē└Ž└ŅĪóŠW░a╔┘─Ļ░óÅŖĄ╚ę╗ŽĄ┴ą─ąąįą╬Ž¾ęį╝░×ķ┤╦│Ņ├╝┐Ó─śĄ─╚²ŗĪó└Ž└ŅŽ▒ŗDā║Īó░óÅŖŗīŗīĪŻį┌╬ęéāĄ─ėĪŽ¾ųąŻ¼¤ošō│ķ¤¤║╚ŠŲ╬³ČŠ▀Ć╩Ū┘Ć▓®┤“ė╬æ“Ż¼─ąąį╦Ų║§Č╝▒╚┼«ąįĖ³╚▌ęū│╔░aŻ¼▀@╩ŪŲ½ęŖ├┤Ż¼▀Ć╩ŪšµīŹ┤µį┌Ą──žŻ┐╚ń╣¹│╔░aĄ─ąįäe▓Ņ«ÉšµĄ─┤µį┌Ż¼▒│║¾Ą─ÖCųŲ╩Ū╩▓├┤Ż┐
2025─Ļ5į┬7╚šŻ¼╚A╩óŅD┤¾īWJose A. MoronĮ╠╩┌łFĻĀį┌ų¬├¹Ų┌┐»NeuronŻ©IF2024=14.7Ż®░l▒Ē┴╦Ņ}├¹×ķ“Estradiol protects against pain-facilitated fentanyl use via suppression of opioid-evoked dopamine activity in males”Ą─蹊┐šō╬─ĪŻčąŠ┐łFĻĀ═©▀^į┌┤¾╩¾╔Ž▀MąąīŹ“ׯ¼“×ūC┴╦│╔░a┤_īŹ┤µį┌ąįäe▓Ņ«ÉŻ¼Č°┤Ų╝ż╦žį┌▀@└’├µ░lō]ų°ųžę¬ū„ė├ĪŻ

蹊┐┴┴³c
1Īó¤oŠĆ╣Ō└wėøõøĄ─ķLŲ┌äėæB▒O£y
īŹ¼F┴╦į┌ūįė╔ąą×ķäė╬’ųąķL▀_öĄų▄Ą─VTAŌ}ą┼╠¢ėøõøŻ¼▓ČūĮĄĮ╠█═┤šTī¦Ą─ČÓ░═░Ę╗ŅäėļSĢrķgųØuį÷ÅŖĄ─äėæB▀^│╠ĪŻ
2Īóąįäe╠ž«ÉąįÖCųŲĄ─Įę╩Š
╩ū┤╬├„┤_čū░Y╠█═┤āHį÷ÅŖą█ąįČ°ĘŪ┤Ųąį┤¾╩¾Ą─Ęę╠½─ß╩╣ė├Ż¼▓ó░l¼FŲõė╔VTAČÓ░═░Ę╔±Įøį¬╗Ņäėį÷ÅŖ“īäėĪŻ
3Īó╝ż╦ž-╠█═┤-╔±Įø┐╔╦▄ąįĄ─╚²ųžĮ╗╗ź
░l¼F┤ŲČ■┤╝Ż©E2Ż®Ą─▒Żūoū„ė├ę└┘ćė┌╠█═┤▒│Š░Ż¼į┌╠█═┤ą█ąįųąęųųŲČÓ░═░Ę╗ŅäėŻ¼Č°į┌¤o╠█═┤┤ŲąįųąĘ┤Č°į÷ÅŖŻ¼╠¶æ┴╦E2å╬╝ā┤┘▀M¬ä┘pĄ─é„Įyė^³cĪŻ
4Īó╗»īW▀zé„īW“×ūCę“╣¹ąį
▀^DREADD╝╝ągūC├„VTAČÓ░═░Ę╔±Įøį¬╗Ņäėī”Ęę╠½─ß╩╣ė├Ą─▒žę¬ąį┼c│õĘųąįĪŻ
5Īó┼R┤▓▐D╗»Øō┴”
╠ß╩Š┤Ų╝ż╦ž╗“ERβ╝żäėä®┐╔─▄ū„×ķ╠█═┤╗╝š▀ųąŅAĘ└░óŲ¼×Eė├Ą─Øōį┌ų╬»¤▓▀┬įŻ¼ė╚Ųõ▀mė├ė┌─ąąį╗“╝ż╦ž╠µ┤·ų╬»¤╚╦╚║ĪŻ
CFAčū░Yą█╩¾▒╚┤Ų╩¾Ė³āAŽ“Ęę╠½─ß╔ŽŅ^
į█éāüĒ┐┤┐┤īŹ“׊▀¾w╩Ūį§├┤ū÷Ą─ĪŻčąŠ┐łFĻĀęį░óŲ¼ŅÉ╦Ä╬’Ęę╠½─ß×ķ┬õ─_³cüĒ蹊┐│╔░aĪŻ░óŲ¼ŅÉ╦Ä╬’ū„×ķÅŖą¦µé═┤╦ÄŻ¼│Żė├ė┌ų╬»¤ųąĄĮųžČ╚╠█═┤ĪŻæł÷╔Ž▒╗š©Ą├╩ų─_ÖM’wĄ─╩┐▒°Ż¼▄ŖßtĢ■×ķ╦¹éā┤“╔Žę╗ßśåßĘ╚üĒµé═┤Ż¼åßĘ╚Š═╩Ūę╗ĘN░óŲ¼ŅÉ╦Ä╬’ĪŻ░óŲ¼ŅÉ╦Ä╬’ļm╩Ūµé═┤Įń┐Ė░čūėŻ¼Ą½┤·ārģs╩ŪŻ¼╗╝š▀┐╔─▄ę“ķLŲ┌╩╣ė├Č°«a╔·╔·└Ēę└┘ć║═─═╩▄ąįŻ¼Å─Č°ī¦ų┬░óŲ¼│╔░aĪŻ┼R┤▓╔Žė^▓ņĄĮŻ¼─ąąį▒╚┼«ąįĖ³ęū×Eė├░óŲ¼ĪŻ
蹊┐łFĻĀ─ŻöM┼R┤▓Ż¼╩ūŽ╚Į©┴ó┴╦╠█═┤Ą─┤¾╩¾─Żą═Ż¼▓ó┼õéõ┴╦ŽÓæ¬Ą─ī”ššĮMŻ¼ą╬│╔4ĮM▓╗═¼Śl╝■Ž┬Ą─┤¾╩¾Ż║─_Ąū░Õūó╔õ═Ļ╚½Ėź╩Įū¶ä®Ż©CFAŻ®Å─Č°«a╔·┬²ąįčū░Y╠█═┤Ą─ą█ąį┤¾╩¾Ż©CFAčū░Yą█╩¾Ż®║═┤Ųąį┤¾╩¾Ż©CFAčū░Y┤Ų╩¾Ż®ęį╝░─_Ąū░Õūó╔õ╔·└Ē¹}╦«Ą─ī”ššą█ąį┤¾╩¾Ż©ī”ššą▄╩¾Ż®║═┤Ųąį┤¾╩¾Ż©ī”šš┤Ų╩¾Ż®ĪŻ
╚╗║¾Įo▀@4ĮM┤¾╩¾Č╝Įė╔Ž┴╦ę╗╠ūčbų├Ż¼ė¢ŠÜ╦³éā▀MąąĘę╠½─ßūį╬ęĮo╦ÄŻ©self administrationŻ¼SAŻ¼łD1Ż®ĪŻčbų├Š▀¾w╩Ū▀@śėĄ─Ż¼┤¾╩¾Ū░├µėąā╔éĆ░┤ŌoŻ¼░┤Ųõųąę╗éĆĢ■åóäėūó╔õŲ„Ą─ķ_ĻPŻ¼═©▀^ņo├}┴¶ų├ßśŽ“┤¾╩¾¾wā╚ūó╔õ100 μLĘę╠½─߯¼▓óŪę░ķļS5 sĄ─╠ß╩Š¤¶┴┴Ż¼░┤┴Ē═Ōę╗éĆ░┤Ōo▓╗Ģ■ėą╚╬║╬Ę┤æ¬ĪŻė¢ŠÜ├┐╠ņ▀Mąą2ąĪĢrŻ¼├┐ų▄5╠ņŻ¼│ų└m3ų▄Ż¼Ą┌1ų▄ūó╔õŲ„└’Ęę╠½─ߥ─ØŌČ╚×ķ5 μg/kgŻ©Ė∙ō■┤¾╩¾Ą─¾wųž┼õų├ØŌČ╚Ż®Ż¼Ą┌2-3ų▄×ķ2 μg/kgĪŻ
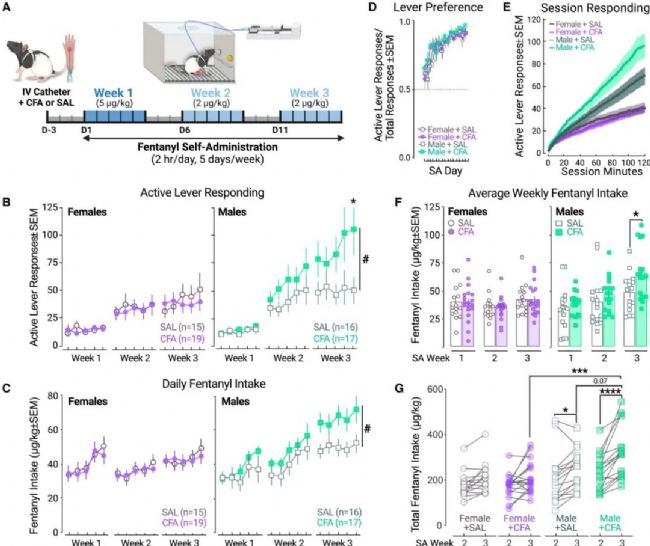
łD 1 čū░Yąį╠█═┤▀xō±ąįĄž┤┘▀Mą█ąį┤¾╩¾Ą─Ęę╠½─ßūį╬ęĮo╦Ä
(A) ąą×ķīŹ“×ĢrķgŠĆ╩ŠęŌłDĪŻ
(B ║═ C) CFA╗“SAL╠Ä└Ē║¾ą█ąį║═┤Ųąį┤¾╩¾Ą─ (B) ėąą¦Ė▄ŚUĘ┤æ¬┤╬öĄ ║═ (C) Įø¾wųžś╦£╩╗»Ą─Ęę╠½─ßöz╚ļ┴┐ĪŻ
(D) ėąą¦┼c¤oą¦Ė▄ŚUĄ─ģ^ĘųŻ©Ė▄ŚUŲ½║├ = ėąą¦Ė▄ŚU░┤ē║┤╬öĄ / ┐éĖ▄ŚU░┤ē║┤╬öĄŻ®ĪŻ
(E) SAĄ┌3ų▄Ų┌ķgŲĮŠ∙ėąą¦Ė▄ŚUĘ┤æ¬╦┘┬╩ĪŻ
(F ║═ G) CFA╗“SAL╠Ä└Ē║¾ą█ąį║═┤Ųąį┤¾╩¾ (F) Ė„SAų▄Ą─ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐ (μg/kg) ęį╝░ (G) SAĄ┌2ų▄ų┴Ą┌3ų▄Ż©2 μg/kg/┤╬Ż®┐éĘę╠½─ßöz╚ļ┴┐Ą─▒╚▌^ĪŻ
(F) ║═ (G) ųąĄ─├┐éĆĘ¹╠¢┤·▒Ēę╗éĆéĆ¾wĪŻ
┐╔ęį┐┤ĄĮŻ¼į┌Ą┌1Īó2ų▄Ż¼4ĮM┤¾╩¾▒Ē¼Fø]ėą▓Ņ«ÉŻ¼Ą┌3ų▄Ż¼CFAčū░Yą█╩¾öz╚ļ┴╦▒╚CFAčū░Y┤Ų╩¾Ė³ČÓĄ─Ęę╠½─ßĪŻ▀@Ą├ĄĮ┴╦Ą┌ę╗éĆĮYšōŻ║čū░Y╠█═┤Ģ■ļSų°ĢrķgĄ─═ŲęŲŻ¼▀xō±ąį┤┘▀Mą█╩¾Ą─Ęę╠½─ßöz╚ļĪŻ
Ęę╠½─ß╔ŽŅ^Ą─ąįäe▓Ņ«ÉĄ─įŁę“╠Į╦„
×ķ╩▓├┤CFAčū░Yą█╩¾Ģ■öz╚ļĖ³ČÓĄ─Ęę╠½─߯┐ļyĄ└ą█╩¾ŽÓ▒╚ė┌┤Ų╩¾Ėą╩▄ĄĮ┴╦Ė³ÅŖ┴ęĄ─═┤ĖąŻ¼╗“š▀═¼ä®┴┐Ą─Ęę╠½─ßį┌ą█╩¾ųąĄ─ą¦╣¹▓╗╚ń┤Ų╩¾Ż¼╦∙ęįąĶę¬öz╚ļĖ³ČÓ┴┐Ą─Ęę╠½─ßüĒ▀_ĄĮ═¼śėĄ─ą¦╣¹Ż┐蹊┐łFĻĀ═©▀^īŹ“×┼┼│²┴╦▀@ā╔éĆ┐╔─▄ąįŻ©łD2Ż®Ż¼▓ó░čĻPūó³c═ČŽ“┴╦Ė╣é╚▒╗╔wģ^Ż©VTAŻ®Ą─ČÓ░═░ĘŻ©DAŻ®╔±Įøį¬ĪŻ
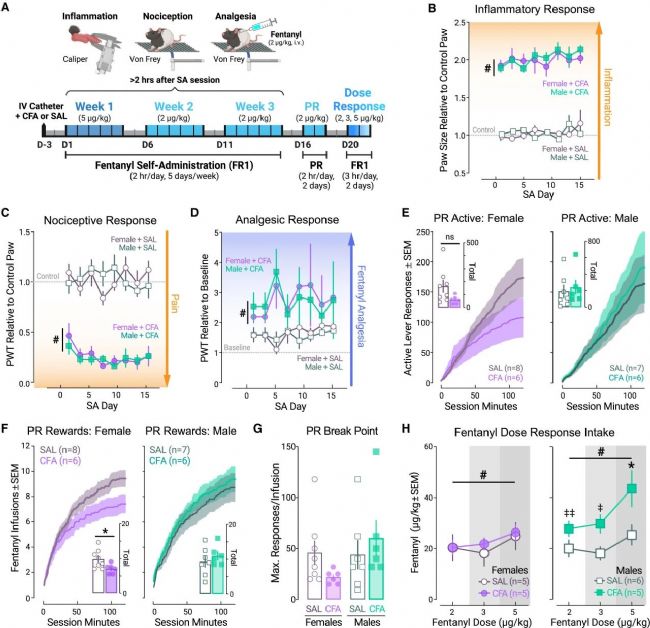
łD 2 CFAį┌ą█ąį║═┤Ųąįųą«a╔·┐╔▒╚╦«ŲĮĄ─čū░YĪóé¹║”ąįĖą╩▄║═Ęę╠½─ßµé═┤ū„ė├Ż¼Ą½į÷ÅŖą█ąįĄ─ä®┴┐Ę┤æ¬
(A) ąą×ķĘĮĘ©╩ŠęŌłDĪŻ
(B) Ęę╠½─ßSA╚²ų▄Ų┌ķgė├£yÅĮŲ„£y┴┐Ą─ŽÓī”ū”║±Č╚ĪŻ
(C) ė├ļŖūėVon Frey£yĄ├Ą─ŽÓī”ū”┐sūŃķōųĄ (PWT)ĪŻ
(D) Ęę╠½─ßšTī¦Ż©2 μg/kg, ņo├}ūó╔õŻ®Ą─µé═┤ū„ė├Ż¼═©▀^▌öūó║¾ŽÓī”ė┌▌öūóŪ░Ą─PWT▒Ē╩ŠĪŻ
(E) ┤ŲąįŻ©ū¾Ż®║═ą█ąįŻ©ė꯮į┌PRŲ┌ķgĄ─ŲĮŠ∙ėąą¦Ė▄ŚUĘ┤æ¬┤╬öĄęį╝░ŲĮŠ∙┐éĘ┤æ¬┤╬öĄŻ©▓ÕłDŻ®ĪŻ
(F) PRļAČ╬½@Ą├Ą─ŲĮŠ∙Ęę╠½─ß▌öūó┤╬öĄŻ©¬ä┘pŻ®ęį╝░½@Ą├Ą─┐é¬ä┘pöĄŻ©▓ÕłDŻ®ĪŻ
(G) ŲĮŠ∙PRųąöÓ³cŻ¼ęį×ķ┴╦ę╗┤╬▌öūó╦∙ū„Ą─ūŅ┤¾Ę┤æ¬┤╬öĄ▒Ē╩ŠĪŻ
(H) ┤ŲąįŻ©ū¾Ż®║═ą█ąįŻ©ė꯮į┌Ęę╠½─ßä®┴┐Ę┤欣yįćųąĄ─ŲĮŠ∙öz╚ļ┴┐ĪŻ
░óŲ¼ŅÉ╦Ä╬’─▄ē“┤╠╝żVTAĄ─DAßīĘ┼Ż¼Č°čąŠ┐łFĻĀų«Ū░Ą─¾w═Ō蹊┐░l¼FŻ¼CFAĢ■ī”VTAĄ─DAßīĘ┼«a╔·ęųųŲū„ė├ĪŻę“┤╦Ż¼░óŲ¼ŅÉ╦Ä╬’ī”ė┌CFA║¾VTAĄ─DAßīĘ┼─▄┴”Ė─ūāūāĄ├ļyęįŅA£yĪŻėąø]ėą┐╔─▄CFAī”VTAĄ─ČÓ░═░Ę╔±Įøį¬Ą─ė░Ēæī¦ų┬┴╦Ęę╠½─ß╩╣ė├ųąĄ─ąįäe▓Ņ«ÉŻ┐蹊┐╚╦åT└¹ė├äōą┬Ą─¤oŠĆ¾wā╚╣Ō└w╣ŌČ╚ėøõø╝╝ągŻ¼īŹĢr▒O£y┴╦Ęę╠½─ßūį╬ęĮo╦Ä▀^│╠ųąVTADA╔±Įøį¬Ą─Ō}╦▓ūā╗ŅäėŻ¼ęį╠ĮŠ┐╠█═┤║═ąįäeę“╦žī”ŲõĄ─ė░ĒæŻ©łD3AŻ®ĪŻ
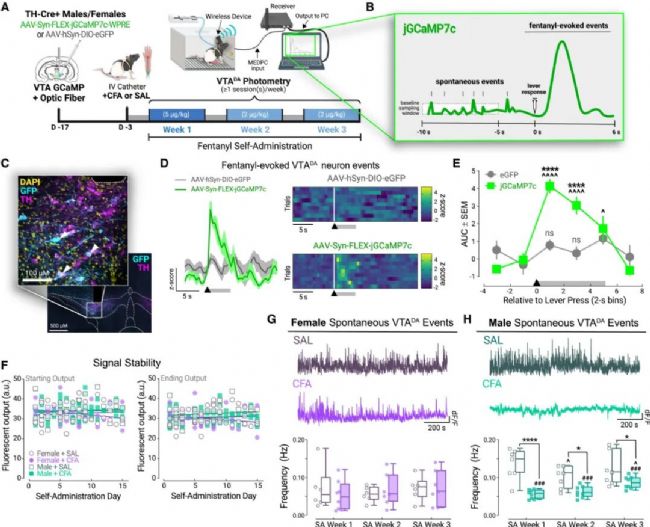
łD 3 Ęę╠½─ßSAŲ┌ķgĄ─¤oŠĆ¾wā╚╣Ō└w╣ŌČ╚ėøõøÖz£yĄĮī”VTADA╔±Įøį¬╗ŅąįĄ─ąįäe║═╠█═┤╠ž«Éąįą¦æ¬
(A) īŹ“×ĘĮĘ©╩ŠęŌłDĪŻ
(B) ūį░lVTADA╩┬╝■║═┼cī¦ų┬Ęę╠½─ß▌öūóĄ─Ė▄ŚU░┤ē║ĢrķgµiČ©Ą─Ęę╠½─ßšT░l╩┬╝■Ą─╩┬╝■Öz£yĪŻ
(C) TH-Cre+┤¾╩¾VTAā╚╣Ō└w╬╗ų├Ż©╠ōŠĆŻ®║═Creę└┘ćĄ─jGCaMP7c (GFP) ▒Ē▀_┼cTHŻ©░ū╔½╝²Ņ^Ż®╣▓Č©╬╗Ą─┤·▒ĒąįłDŽ±ĪŻ
(D) ▒Ē▀_jGCaMP7c╗“ī”šš▓ĪČŠĄ─┤¾╩¾ųąŻ¼Ęę╠½─ßšT░lĄ─VTADA╔±Įøį¬¤╔╣ŌĄ─ZųĄęį╝░å╬éĆįć“ץ─¤ßłDŻ¼▀@ą®įć“×┼cėąą¦Ė▄ŚU░┤ē║Ż©║┌╔½╝²Ņ^Ż®║═Ęę╠½─ß▌öūó/ŠĆ╦„│╩¼FŻ©╗ę╔½ŚlŻ®ĢrķgµiČ©ĪŻ▒╚└²│▀Ż║1 ZųĄŻ©┤╣ų▒Ż®║═ 5├ļŻ©╦«ŲĮŻ®ĪŻ
(E) ▒Ē▀_jGCaMP7c╗“ī”šš▓ĪČŠĄ─VTADA╔±Įøį¬ųąŻ¼Ęę╠½─ßšT░l╩┬╝■ęį2├ļķgĖ¶ĘųĮMĄ─Ū·ŠĆŽ┬├µĘe (AUC)ĪŻ
(F) SAĢ■įÆķ_╩╝╗“ĮY╩°ĢrĄ─¤╔╣ŌŻ©╚╬ęŌå╬╬╗, a.u.Ż®ų«ķgĄ─ĻPŽĄĪŻöĄō■³c┤·▒Ēå╬éĆėøõøĢ■įÆĪŻ
(G ║═ H) (G) CFA╗“SAL╠Ä└ĒĄ─┤Ųąį║═ (H) ą█ąįĄ─ūį░lVTADAŌ}╦▓ūā╗Ņąį┤·▒Ēąį▄ē█EŻ©╔ŽłDŻ®║═ŲĮŠ∙├┐ų▄ūį░l╩┬╝■┬╩Ņl┬╩Ż©Ž┬łDŻ®ĪŻöĄō■³c┤·▒ĒüĒūįå╬éĆ┤¾╩¾Ą─├┐ų▄Š∙ųĄĪŻ
ĮY╣¹’@╩ŠŻ¼CFAčū░Y╠█═┤ī”VTADA╔±Įøį¬Ą─╗∙ĄAūį░l╗Ņäė«a╔·ąįäe╠ž«ÉĄ─š{┐žŻ║╦³│ų└mĮĄĄ═┴╦ą█ąį┤¾╩¾Ą─ūį░l╗ŅäėŅl┬╩Ż¼Č°ī”┤Ųąįät¤oė░ĒæŻ©łD3G,HŻ®ĪŻčąŠ┐╚╦åT│╔╣”ģ^Ęų┴╦ūį░lĄ─╔±Įø╗Ņäė┼c“Ęę╠½─ßšT░l”Ą─╗ŅäėŻ©ė╔Ė▄ŚU░┤ē║ĪóŠĆ╦„╠ß╩Š║═╦Ä╬’▌öūó╣▓═¼ė|░lŻ®ĪŻĘę╠½─ßšT░l╗ŅäėĄ─«a╔·ąĶę¬Ė▄ŚU░┤ē║Ę┤æ¬ ĪóŠĆ╦„│╩¼Fęį╝░═¼ĢrĄ─Ęę╠½─ß▌ö╦═Ż¼▀@ą®ę“╦ž»B╝ėŻ¼╣▓═¼ę²Ų┴╦VTADA╔±Įøį¬Ō}╦▓ūā╗ŅäėĄ─╝▒äĪį÷╝ėŻ©łD3BŻ¼ęĢŅl1Īó2Ż®ĪŻ
ęĢŅl1 ėøõøĄĮę╗┤╬“Ęę╠½─ßšT░l”Ą─╗Ņäė

ęĢŅl2 ėøõøĄĮ┴Ēę╗┤╬“Ęę╠½─ßšT░l”Ą─╗Ņäė
蹊┐╚╦åT▀Mę╗▓Į╠Į╦„┴╦SAŲ┌ķgVTADA╔±Įøį¬╗Ņäė╚ń║╬╩▄ĄĮCFAĄ─ė░ĒæŻ©łD4Ż®ĪŻį┌Ęę╠½─ßSA▀^│╠ųąŻ¼┤Ųąį┤¾╩¾Ą─╔±ĮøĘ┤æ¬╩╝ĮK▒Ż│ųĘĆČ©Ż¼▓╗╩▄╠█═┤ė░ĒæĪŻČ°CFAą█ąį┤¾╩¾│§Ų┌Ę┤欣p╚§Ż¼Ą½ļSų°ė├╦ÄĢrķg═ŲęŲŻ¼ŲõĘę╠½─ßšT░lĄ─ČÓ░═░Ę╔±Įøį¬╗ŅäėĘ∙Č╚║═Ę┤æ¬Ė┼┬╩ųų▄’@ų°į÷ÅŖŻ¼į┌Ą┌╚²ų▄▀_ĄĮūŅĖ▀╦«ŲĮŻ¼’@ų°│¼▀^╦∙ėąŲõ╦¹ĮMĪŻ▀@ĘN╔±Įø╗ŅäėĄ─į÷ÅŖ▄ē█E┼cCFAą█ąį┤¾╩¾ąą×ķ╔Ž▒Ē¼F│÷Ęę╠½─ßöz╚ļ┴┐į÷╝ėĄ──Ż╩Į═Ļ╚½ŲĮąąŻ¼Ūę╔±ĮøĘ┤æ¬Ą─ĘÕųĄ┤¾ąĪ┼c╦Ä╬’öz╚ļ┐é┴┐│╩ÅŖš²ŽÓĻPĪŻ
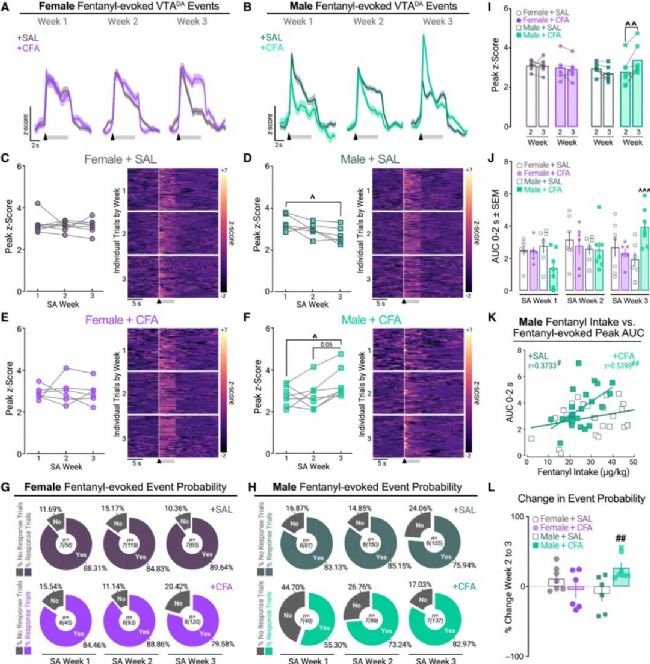
łD 4 ╠█═┤ļSĢrķgį÷╝ėą█ąį┤¾╩¾Ęę╠½─ßšT░lĄ─VTADA╔±Įøį¬╩┬╝■Ę∙Č╚║═Ė┼┬╩
(A ║═ B) (A) ėą╠█═┤Ż©CFA; n=6Ż®╗“¤o╠█═┤Ż©SAL; n=7Ż®Ą─┤Ųąį║═ (B) ėą╠█═┤Ż©CFA; n=7Ż®╗“¤o╠█═┤Ż©SAL; n=6Ż®Ą─ą█ąįųąŻ¼Ęę╠½─ßšT░lĄ─VTADA╩┬╝■ (ZųĄ ± SEM)Ż¼┼cSA├┐ų▄Ą─ėąą¦Ė▄ŚUĘ┤æ¬Ż©║┌╔½╝²Ņ^Ż®║═Ęę╠½─ß▌öūóŻ©╗ę╔½ŚlŻ®ī”²RĪŻ▒╚└²│▀Ż║1 ZųĄŻ©┤╣ų▒Ż®║═ 2├ļŻ©╦«ŲĮŻ®ĪŻ
(C–H) Ė„SAų▄Ą─╩┬╝■ĘÕųĄZųĄęį╝░’@╩Šå╬éĆįć“ץ─¤ßłDŻ¼▀@ą®įć“×┼cī¦ų┬▌öūó/ŠĆ╦„é„▀fĄ─Ė▄ŚU░┤ē║ī”²RŻ¼Ęųäeī”æ¬ (C) ¤o╠█═┤┤ŲąįĪó(D) ¤o╠█═┤ą█ąįĪó(E) ėą╠█═┤┤Ųąį ║═ (F) ėą╠█═┤ą█ąįĪŻ(G) SAL╗“CFA╠Ä└Ē║¾┤Ųąį║═ (H) SAL╗“CFA╠Ä└Ē║¾ą█ąįųąŻ¼├┐┤╬Ęę╠½─ßĖ▄ŚU░┤ē║ę²░l┐╔Öz£yĘÕųĄĄ─Ė┼┬╩ĪŻ
(I) SAĄ┌2ų▄║═Ą┌3ų▄ų«ķgĄ─ĘÕųĄZųĄĪŻ
(J) SA├┐ų▄Ą─VTADA╗ŅąįAUCĪŻ├┐éĆöĄō■³c┤·▒Ēå╬éĆ┤¾╩¾/SAų▄Ą─Š∙ųĄĪŻ
(K) SAL║═CFA╠Ä└Ē║¾ą█ąįųąŻ¼ŲĮŠ∙Ęę╠½─ßšT░lVTADA AUC┼cSAŲ┌ķgĘę╠½─ßöz╚ļ┴┐ų«ķgĄ─ĻP┬ōĪŻ
(L) SAĄ┌2ų▄ų┴Ą┌3ų▄VTADAĘÕųĄĖ┼┬╩Ą─░┘Ęų▒╚ūā╗»ĪŻ
į÷ÅŖĄ─VTADA╔±Įøį¬╗Ņäė┤┘╩╣Ęę╠½─ß╔ŽŅ^
×ķ┤_Č©Ė▀ÅŖČ╚VTAČÓ░═░Ę╔±Įøį¬╗Ņäė╩Ūʱ×ķCFAą█ąį┤¾╩¾į÷╝ėĘę╠½─ß╩╣ė├╦∙▒žąĶŻ¼čąŠ┐╚╦åTį┌SAĄ┌╚²ų▄▓╔ė├╗»īW▀zé„īWĘĮĘ©ęųųŲVTADA╔±Įøį¬Ż¼▓óÖz£yŲõī”Ęę╠½─ßšT░lĄ─VTADA╗Ņäė╝░Ęę╠½─ßöz╚ļ┴┐Ą─ė░ĒæŻ©łD5Ż®ĪŻīŹ“×░l¼FŻ¼╩╣ė├Gi-DREADDęųųŲ╔±Įøį¬║¾Ż¼įŁ▒Šį┌Ą┌╚²ų▄Ģ■│÷¼FĄ─Ęę╠½─ßöz╚ļ┴┐į÷ķL▒╗═Ļ╚½ūĶöÓŻ¼Ųõöz╚ļ╦«ŲĮŠS│ųį┌Ą┌Č■ų▄Ą─│╠Č╚Ż¼Ūęį┌äėÖC£yįćųąĄ─Ę┤æ¬ę▓£p╔┘ĪŻ═¼ĢrŻ¼Ęę╠½─ßšT░lĄ─DA╔±Įø╗Ņäėę▓ŽÓ欎┬ĮĄĪŻĮY╣¹▒Ē├„Ż¼╠█═┤▒│Š░Ž┬ą█ąį┤¾╩¾Ęę╠½─ß╩╣ė├┴┐Ą─į÷╝ėŻ¼ę└┘ćė┌VTADA╔±Įøį¬ī”╦Ä╬’Ę┤æ¬ąįļSĢrķg═ŲęŲČ°į÷ÅŖĄ─▀@ę╗╠žČ©╔±Įø╗Ņäėūā╗»Ż¼ūC├„┴╦įō╔±ĮøÖCųŲĄ─ąą×ķ▒žę¬ąįĪŻ
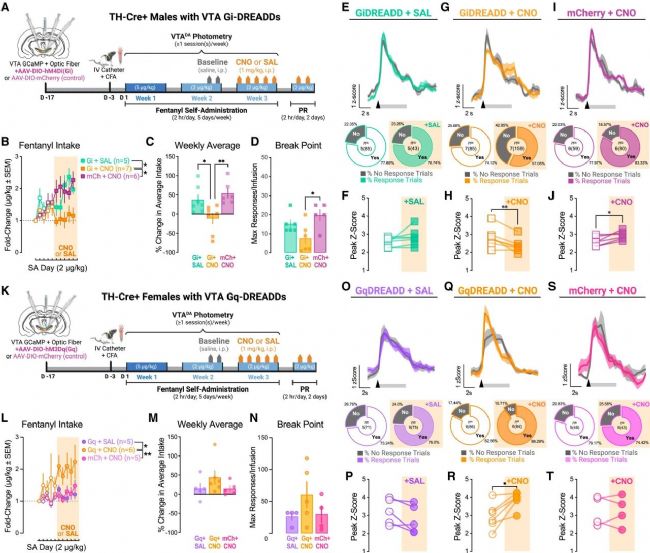
łD 5 SAĄ┌3ų▄Ų┌ķgĖ▀ÅŖČ╚Ęę╠½─ßšT░lVTADA╗Ņąį╩ŪCFA║¾┤┘▀MĘę╠½─ß╩╣ė├Ą─▒žę¬║═│õĘųŚl╝■
(A) ą█ąįīŹ“×ĘĮĘ©╩ŠęŌłDĪŻSAĄ┌3ų▄Ų┌ķgŻ¼▒Ē▀_Gi-DREADDĄ─ą█ąįį┌SAĢ■įÆķ_╩╝Ū░20ĘųńŖĮė╩▄SAL (Gi+SAL) ╗“CNO (Gi+CNO)Ż¼▒Ē▀_mCherryĄ─ą█ąįĮė╩▄CNO (mCh+CNO)ĪŻ
(B) CNO╗“SAL╠Ä└Ē║¾Ż¼ą█ąįį┌Ęę╠½─ßSAŻ©2 μg/kg/┤╬Ż®Ų┌ķg├┐╚šĘę╠½─ßöz╚ļ┴┐Ą─ūā╗»ĪŻ
(C) ┼c╗∙ŠĆŽÓ▒╚Ż¼ą█ąįį┌CNO╗“SAL║¾Ą─ŲĮŠ∙öz╚ļ┴┐ūā╗»ĪŻ
(D) CNO╗“SAL║¾ą█ąįĄ─PRųąöÓ³cĪŻ(E ║═ F) Gi+SALĪó(G ║═ H) Gi+CNO ║═ (I ║═ J) mCh+CNO ą█ąįųąŻ¼┼cĖ▄ŚU░┤ē║Ż©╝²Ņ^Ż®║═▌öūóŻ©╗ę╔½ŚlŻ®ī”²RĄ─Ęę╠½─ßšT░lVTADAŌ}╦▓ūāĪóĘÕųĄĖ┼┬╩║═ĘÕųĄZųĄŻ©╗∙ŠĆĢrŻ®ĪŻ
(K) ┤ŲąįīŹ“×ĘĮĘ©╩ŠęŌłDĪŻSAĄ┌3ų▄Ų┌ķgŻ¼▒Ē▀_Gq-DREADDĄ─┤Ųąįį┌SAĢ■įÆķ_╩╝Ū░20ĘųńŖĮė╩▄SAL (Gq+SAL) ╗“CNO (Gq+CNO)Ż¼▒Ē▀_mCherryĄ─┤ŲąįĮė╩▄CNO (mCh+CNO)ĪŻ
(L) CNO╗“SAL╠Ä└Ē║¾Ż¼┤Ųąįį┌Ęę╠½─ßSAŻ©2 μg/kg/┤╬Ż®Ų┌ķg├┐╚šĘę╠½─ßöz╚ļ┴┐Ą─ūā╗»ĪŻ
(M) ┼c╗∙ŠĆŽÓ▒╚Ż¼┤Ųąįį┌CNO╗“SAL║¾Ą─ŲĮŠ∙öz╚ļ┴┐ūā╗»ĪŻ
(N) CNO╗“SAL║¾┤ŲąįĄ─PRųąöÓ³cĪŻ(O ║═ P) Gq + SALĪó(Q ║═ R) Gq + CNO ║═ (S ║═ T) mCh + CNO ┤ŲąįųąŻ¼┼cĖ▄ŚU░┤ē║║═▌öūóī”²RĄ─Ęę╠½─ßšT░lVTADAŌ}╦▓ūāĪóĘÕųĄĖ┼┬╩║═ĘÕųĄZųĄĪŻ
蹊┐╚╦åT▀Mę╗▓Į═©▀^╗»īW▀zé„īW╝╝ąg╝ż╗ŅCFA┤Ųąį┤¾╩¾Ą─VTAČÓ░═░Ę╔±Įøį¬Ż¼░l¼F▀@ĘN╚╦×ķĄ─┼dŖ^ąįĖ╔ŅAūŃęį─ŻöMą█ąį▒Ēą═Ż¼ī¦ų┬ŲõĘę╠½─ßöz╚ļ┴┐╝░╦Ä╬’šT░lĄ─DA╔±Įøį¬Ę┤欒@ų°į÷╝ėŻ©łD5Ż®ĪŻįōĮY╣¹┼cęųųŲīŹ“׎ÓĮY║ŽŻ¼╣▓═¼ūC├„┴╦į÷ÅŖĄ─VTADA╔±Įøį¬╗Ņäė╩Ū“īäė╠█═┤▒│Š░Ž┬Ęę╠½─ß╩╣ė├┴┐į÷╝ėĄ─│õĘų▒žę¬Śl╝■Ż¼Įę╩Š┴╦ČÓ░═░ʬä┘pŽĄĮy▀^Č╚┼dŖ^į┌│╔░aąą×ķųąĄ─║╦ą─ę“╣¹ū„ė├ĪŻ
┤ŲČ■┤╝Ż©E2Ż®ī¦ų┬Ą─ąįäe▓Ņ«É
ė╔ė┌╠█═┤Ģ■┤┘▀MĘę╠½─ßšT░lĄ─VTADA╔±Įøį¬Ę┤æ¬Ż¼Å─Č°▀xō±ąįĄž“īäėą█ąįį÷╝ėĘę╠½─ß╩╣ė├Ż¼čąŠ┐╚╦åT═Ų£yŽ╚╠ņĄ─ąįäe▓Ņ«É▒Żūo┴╦┤ŲąįĪŻ═©▀^▀Mę╗▓Į╠Į╦„Ż¼čąŠ┐╚╦åT░l¼F┤Ųąį┤¾╩¾ī”Ęę╠½─ߥ─╠ņ╚╗Ąų┐╣┴”▓óĘŪė╔äėŪķų▄Ų┌š{╣ØŻ¼Č°╩Ūę└┘ćė┌┬č│▓╝ż╦žĄ─▒Żūoū„ė├Ż©łD6Ż®ĪŻ╚ź│²┬č│▓Ż©OVXŻ®║¾Ż¼┤Ųąį┤¾╩¾Ą─Ęę╠½─ßöz╚ļ┴┐╝░ŲõšT░lĄ─VTAų┴Ę³Ė¶║╦Ą─ČÓ░═░Ę╔±Įø═©┬Ę╗ŅäėŠ∙’@ų°į÷ÅŖŻ¼Ųõ▒Ē¼F┼c╠█═┤ĀŅæBŽ┬Ą─ą█ąį┤¾╩¾ŽÓ╦ŲĪŻ▀@▒Ē├„┬č│▓Ęų├┌Ą─╝ż╦žŻ©╚ń┤Ų╝ż╦žŻ®─▄ėąą¦ęųųŲČÓ░═░ʬä┘pŽĄĮyī”░óŲ¼╦Ä╬’Ą─Ę┤æ¬ąįŻ¼Å─Č°į┌╔·└ĒĀŅæBŽ┬ęųųŲ┤ŲąįĄ─╦Ä╬’īżŪ¾ąą×ķŻ¼śŗ│╔┴╦ąįäe▓Ņ«ÉĄ─ĻPµIā╚į┌▒ŻūoÖCųŲĪŻ
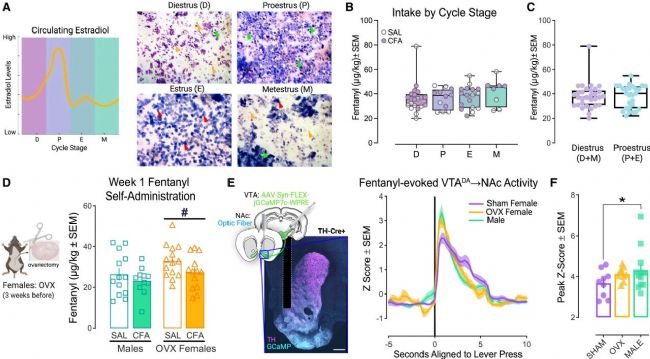
łD 6 ┬č│▓╝ż╦žĄ─║─Į▀┤┘▀MĘę╠½─ßūį╬ęĮo╦Ä▓óęū╗»Ęę╠½─ßšT░lĄ─VTADA→NAc╗Ņąį
(A) ╦─éĆų▄Ų┌ļAČ╬——äėŪķķgŲ┌ (D)ĪóäėŪķŪ░Ų┌ (P)ĪóäėŪķŲ┌ (E)ĪóäėŪķ║¾Ų┌ (M)——ŽÓī”┤Ų╝ż╦ž╦«ŲĮ╩ŠęŌłDŻ©ū¾Ż®ęį╝░░³║¼░ū╝Ü░¹Ż©³S╔½╝²Ņ^Ż®Īóėą║╦╔ŽŲż╝Ü░¹Ż©ŠG╔½╝²Ņ^Ż®║═ĮŪ┘|╗»╔ŽŲż╝Ü░¹Ż©╝t╔½╝²Ņ^Ż®Ą─┤·▒ĒąįĻÄĄ└╝Ü░¹īW═┐Ų¼ĪŻ
(B) SAĄ┌2-3ų▄Ż©2 μg/kg/┤╬Ż®Ė„ļAČ╬Ą─ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐ĪŻ
(C) ┤ŲČ■┤╝╦«ŲĮĄ═ (D+M) ╗“Ė▀ (P+E) ĢrĄ─ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐ĪŻ
(D) ┬č│▓Ūą│²ąg (OVX) │╠ą“╩ŠęŌłD║═Ęę╠½─ßūį╬ęĮo╦ÄĄ┌1ų▄Ų┌ķgĄ─ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐ĪŻ
(E) ▓ĪČŠ░ąŽ“▓▀┬į╩ŠęŌłDęį╝░jGCaMP┼cTH╣▓Č©╬╗║═╣Ō└w╬╗ų├Ą─┤·▒ĒąįłDŽ±Ż©ū¾Ż╗▒╚└²│▀Ż║500 μmŻ®Ż╗ą█ąįŻ©╦{ŠG╔½Ż®ĪóOVX┤ŲąįŻ©Į╔½Ż®╗“╝┘╩ųągOVX (SHAM; ūŽ╔½) Ą─ąįŽ┘═Ļš¹┤ŲąįųąĘę╠½─ßšT░lĄ─VTADA→NAc╦▓ūāĪŻ
(F) ╠█═┤ĀŅæBŽ┬Ż¼SHAM╗“OVX╠Ä└ĒĄ─┤Ųąį╝░═Ļš¹ą█ąįųąŻ¼Ęę╠½─ßšT░lĄ─VTADA→NAc╦▓ūāĄ─ĘÕųĄZųĄĪŻ
┤ŲČ■┤╝Ż©E2Ż®į┌┤Ųąį¾wā╚ØŌČ╚▌^Ė▀Ż¼▓ó┐╔š{╣ØČÓ░═░ĘŻ©DAŻ®╗Ņąį║═¬ä┘pŽÓĻPąą×ķĪŻ×ķ┤_Č©E2╩ŪʱęųųŲ╠█═┤┤┘▀MĄ─Ęę╠½─ß╩╣ė├║═VTADA╗ŅąįŻ¼į┌SAĄ┌╚²ų▄Ž“OVX┤Ųąį║══Ļš¹ą█ąį╚½╔ĒąįĮoėĶE2Ż©łD7Ż®ĪŻčąŠ┐░l¼F┤ŲČ■┤╝Ż©E2Ż®ī”Ęę╠½─ß╩╣ė├╝░ŲõšT░lVTAČÓ░═░Ę╔±Įøį¬╗ŅäėĄ─š{╣Øū„ė├Š▀ėą’@ų°Ą─“ŪķŠ│ę└┘ćąį”Ż¼╝┤═¼Ģr╚ĪøQė┌ąįäe║═╠█═┤ĀŅæBĪŻį┌╠█═┤▒│Š░Ž┬Ą─ą█ąį┤¾╩¾ųąŻ¼E2░lō]┴╦▒Żūoąįū„ė├Ż¼’@ų°ęųųŲ┴╦Ęę╠½─ßšT░lĄ─DA╔±Įøį¬╗Ņäė╝░╦Ä╬’öz╚ļ┴┐ĪŻ╚╗Č°Ż¼į┌OVXĄ─┤ŲąįųąŻ¼E2Ą─ą¦╣¹Įž╚╗ŽÓĘ┤Ż║į┌¤o╠█═┤ĀŅæBŽ┬Ż¼╦³Ę┤Č°į÷ÅŖ┴╦ČÓ░═░ĘĘ┤æ¬║═╦Ä╬’öz╚ļŻ╗Č°į┌╠█═┤┤µį┌ĢrŻ¼E2ät╩¦╚źš{╣Øū„ė├ĪŻ▀@▒Ē├„E2Ą─╔±Įø╦Ä└ĒīWą¦æ¬▓óĘŪ╣╠Č©▓╗ūāŻ¼ŲõĘĮŽ“Ż©ęųųŲ╗“┤┘▀MŻ®ć└ųžę└┘ćė┌ÖC¾wā╚į┌Ą─╝ż╦ž▒│Š░║══Ōį┌Ą─╠█═┤ĀŅæBŻ¼Įę╩Š┴╦╝ż╦ž-ŁhŠ│╗źū„į┌š{┐ž│╔░aęūĖąąįųąĄ─Å═ļsąįĪŻ

łD 7 ┤ŲČ■┤╝ī”Ęę╠½─ßSA║═VTADA╗ŅąįĄ─ą¦æ¬╚ĪøQė┌╠█═┤Ą─┤µį┌
(A) īŹ“×ĘĮĘ©╩ŠęŌłDĪŻOVX┤Ųąį║═ą█ąįį┌ķ_╩╝SAŪ░Įė╩▄CFA╗“SAL╝░ņo├}ī¦╣▄ĪŻĄ┌3ų▄Ģ■įÆŪ░30ĘųńŖŻ¼ŲżŽ┬ūó╔õ┤ŲČ■┤╝ (E2; 20 μg/kg) ╗“▌d¾w (VEH; ųź┬ķė═)ĪŻ
(B ║═ C) CFAą█ąįį┌E2╗“VEH╠Ä└Ē║¾Ż¼SAĄ┌3ų▄Ų┌ķg (B) ┼cĖ▄ŚU░┤ē║Ż©╝²Ņ^Ż®║═Ęę╠½─ß▌öūóŻ©╗ę╔½ŚlŻ®ī”²RĄ─Ęę╠½─ßšT░lVTADAŌ}╦▓ūā╗Ņąį ęį╝░ (C) ĘÕųĄZųĄĪŻ
(D ║═ E) SALą█ąįį┌ (D) E2╗“VEH╠Ä└Ē║¾Ą─VTADAŌ}╦▓ūā╗Ņąį ęį╝░ (E) ĘÕųĄZųĄĪŻ
(F ║═ G) CFA OVX┤Ųąįį┌ (F) E2╗“VEH╠Ä└Ē║¾Ą─VTADA╗Ņąį ęį╝░ (G) ĘÕųĄZųĄĪŻ
(H–M) SAL OVX┤Ųąįį┌ (H) E2╗“VEH╠Ä└Ē║¾Ą─VTADA╗Ņąį ęį╝░ (I) ĘÕųĄZųĄĪŻĘę╠½─ßöz╚ļ┴┐Ą─ūā╗»Ż║(J) CFAą█ąįį┌E2╗“VEH║¾Ż¼(K) CFA OVX┤Ųąįį┌E2╗“VEH║¾Ż¼(L) SALą█ąįŻ¼(M) SAL OVX┤ŲąįĪŻ
(N ║═ P) CFA║═E2╗“VEH╠Ä└Ē║¾ą█ąį║═OVX┤ŲąįĄ─ (N) ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐Ż©2 μg/kg/┤╬Ż®Īó(O) SAĄ┌3ų▄Ų┌ķgŲĮŠ∙öz╚ļ┴┐ ║═ (P) PRųąöÓ³cĪŻ
(Q–S) SAL║═E2╗“VEH╠Ä└Ē║¾ą█ąį║═OVX┤ŲąįĄ─ (Q) ŲĮŠ∙Ęę╠½─ßöz╚ļ┴┐Ż©2 μg/kg/┤╬Ż®Īó(R) SAĄ┌3ų▄Ų┌ķgŲĮŠ∙öz╚ļ┴┐ ║═ (S) PRųąöÓ³cĪŻ
蹊┐╚╦åT▀Mę╗▓Į╠ĮŠ┐┴╦E2ęųųŲ╠█═┤▒│Š░Ž┬Ęę╠½─ß╩╣ė├Ą─ĘųūėÖCųŲŻ¼░l¼FŲõ═©▀^ū„ė├ė┌VTAā╚Ą─┤Ų╝ż╦ž╩▄¾wβŻ©ERβŻ®üåą═░lō]▒Żūoū„ė├Ż©łD8Ż®ĪŻį┌ąįŽ┘═Ļš¹Ą─CFAą█ąį┤¾╩¾ųąŻ¼¤ošō╩Ū╚½╔Ē▀Ć╩ŪVTAŠų▓┐ĮoėĶE2Ż¼Š∙─▄’@ų°ęųųŲĘę╠½─ßšT░lĄ─VTAų┴NAcČÓ░═░Ę═©┬Ę╗ŅäėĪŻĻPµIĄ─╩ŪŻ¼▀@ĘNęųųŲą¦æ¬┐╔▒╗VTAā╚ūó╔õERβ╠ž«Éąį▐ū┐╣䮯©PHTPPŻ®╦∙─µ▐DŻ¼╗ųÅ═Ė▀ÅŖČ╚Ą─╔±ĮøĘ┤æ¬Ż¼Č°ERα▐ū┐╣䮯©MPPŻ®ät¤oą¦Ż¼▒Ē├„E2Ą─▒Żūoū„ė├╠ž«Éąįė╔ERβĮķī¦ĪŻ▀@ĘNERβĄ─š{┐žŠ▀ėąć└Ė±Ą─▒│Š░ę└┘ćąįŻ║╦³āHį┌ąįŽ┘═Ļš¹Īó┤µį┌ā╚į┤ąį╗“═Ōį┤ąį▌^Ė▀╦«ŲĮE2Ą─éĆ¾wųąŲū„ė├ĪŻį┌┬č│▓Ūą│²Ż©OVXŻ®Ą─┤ŲąįųąŻ¼E2╩¦╚ź┴╦š{╣Øū„ė├Ż¼ER▐ū┐╣ä®ę▓¤oą¦Ż¼šf├„═Ļš¹Ą─┬č│▓╝ż╦žŁhŠ│╩ŪERβ╣”─▄Ą─╗∙ĄAĪŻČ°į┌╝┘╩ųągŻ©SHAMŻ®Ą─═Ļš¹┤ŲąįųąŻ¼ERβ▐ū┐╣ät─▄Ž±į┌ą█ąįųąę╗śėį÷ÅŖ╔±ĮøĘ┤æ¬ĪŻ▀@ą®ĮY╣¹Įę╩Š┴╦╠█═┤═©▀^╝ż╗ŅVTA ERβą┼╠¢═©┬ĘŻ¼ąįäe╠ž«ÉąįĄžį÷ÅŖČÓ░═░ĘŽĄĮyī”░óŲ¼╦Ä╬’Ą─Ę┤æ¬ąįŻ¼Å─Č°┤┘▀M│╔░aąą×ķŻ¼×ķķ_░l░ąŽ“ERβĄ─ąįäe╠ž«Éąį│╔░aĖ╔ŅA▓▀┬į╠ß╣®┴╦ĻPµI└Ēšōę└ō■ĪŻ
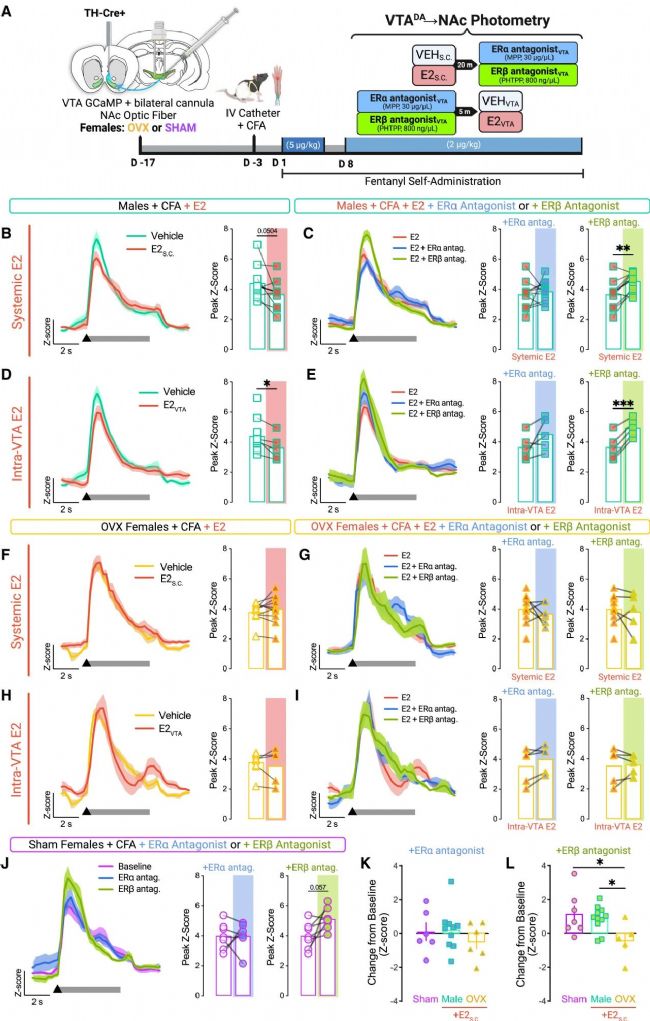
łD 8 ┤ŲČ■┤╝═©▀^VTA┤Ų╝ż╦ž╩▄¾wβą┼╠¢ęųųŲĘę╠½─ßšT░lĄ─VTADA→NAc╗Ņąį
(A) īŹ“×│╠ą“╩ŠęŌłDĪŻTH-Cre+ą█ąį║═┤Ųąį┤¾╩¾į┌VTAĮė╩▄jGCaMP7c▓óų▓╚ļļpé╚╠ū╣▄Ż¼═¼Ģrį┌NAcų▓╚ļå╬é╚╣Ō└wĪŻ┤ŲąįĮė╩▄OVX╗“SHAM╩ųągĪŻ┤¾╩¾į┌Ęę╠½─ßSAŪ░Įė╩▄ņo├}ī¦╣▄║═CFAĪŻį┌╚½╔Ēąįūó╔õE2s.c. (20 μg/kg) ╗“VEHs.c. Ż©ųź┬ķė═Ż╗1 mL/kgŻ®20ĘųńŖ║¾Ż¼┤¾╩¾Įė╩▄VTAā╚ER▐ū┐╣䮥─╬ó┴┐ūó╔õĪŻ╗“š▀Ż¼┤¾╩¾į┌VTAā╚ūó╔õE2VTA (40 ng/μL) ╗“VEHVTA Ż©10% EtOH╔·└Ē¹}╦«Ż®Ū░5ĘųńŖĮė╩▄VTAā╚ER▐ū┐╣ä®ūó╔õĪŻ
(B ║═ C) CFAą█ąįį┌ (B) ╚½╔ĒąįE2s.c. ╗“VEHs.c. ║¾ ęį╝░ (C) E2s.c. ║═MPPVTA ╗“PHTPPVTA ║¾Ą─Ęę╠½─ßšT░lVTADA→NAc╗Ņąį║═ĘÕųĄZųĄĪŻ
(D ║═ E) CFAą█ąįį┌ (D) E2VTA ╗“VEHVTA ║¾ ęį╝░ (E) E2VTA Ū░ŅA╠Ä└ĒMPPVTA ╗“PHTPPVTA ║¾Ą─Ęę╠½─ßšT░lVTADA→NAc╗ŅąįĪŻ
(F ║═ G) CFA OVX┤Ųąįį┌ (F) ╚½╔ĒąįE2s.c. ╗“VEHs.c. ║¾ ęį╝░ (G) E2s.c. ║═MPPVTA ╗“PHTPPVTA ║¾Ą─Ęę╠½─ßšT░lVTADA→NAc╗Ņąį║═ĘÕųĄZųĄĪŻ
(H ║═ I) CFA OVX┤Ųąįį┌ (H) E2VTA ╗“VEHVTA ║¾ ęį╝░ (I) E2VTA Ū░ŅA╠Ä└ĒMPPVTA ╗“PHTPPVTA ║¾Ą─Ęę╠½─ßšT░lVTADA→NAc╗ŅąįĪŻ
(J–L) CFA SHAM┤Ųąįį┌ (J) MPPVTA ╗“PHTPPVTA ║¾Ą─Ęę╠½─ßšT░lVTADA→NAc╗ŅąįĪŻVTAā╚ER▐ū┐╣ä® (K) MPPVTA ╗“ (L) PHTPPVTA ╠Ä└Ē║¾ĘÕųĄZųĄūā╗»Ą─▒╚▌^ĪŻ
蹊┐ĮYšō
蹊┐╩ū┤╬Įę╩Š┬²ąį╠█═┤═©▀^ąįäe╠ž«ÉąįÖCųŲ“īäė░óŲ¼│╔░aŻ║į┌ą█ąįųąŻ¼╠█═┤ę└┘ćąįĄžį÷ÅŖVTAČÓ░═░Ę╔±Įøį¬ī”Ęę╠½─ߥ─Ę┤æ¬ąįŻ¼ų▒Įėī¦ų┬ė├╦Ä┴┐į÷╝ėŻ╗Č°┤Ųąįį┌┬č│▓╝ż╦žŻ©ų„ę¬═©▀^VTA ERβ╩▄¾wŻ®▒ŻūoŽ┬├Ō╩▄┤╦ą¦æ¬ĪŻ
įō░l¼F▓╗āHĻU├„┴╦╠█═┤-│╔░a╣▓▓ĪĄ─╔±Įø╔·╬’īW╗∙ĄAŻ¼Ė³×ķķ_░ląįäe╠ž«ÉąįĄ─Š½£╩µé═┤┼cĘ└Å═╬³▓▀┬į╠ß╣®┴╦Ė’├³ąį░ą³cĪŻ
ģó┐╝╬─½IŻ║
Higginbotham JA, Abt JG, Teich RH, et al. Estradiol protects against pain-facilitated fentanyl use via suppression of opioid-evoked dopamine activity in males. Neuron. 2025 May 7;113(9):1413-1429.e5. doi: 10.1016/j.neuron.2025.02.013.
Žļę¬½@╚Ī▒ŠŲ¬╬─½IĄ─└ŽÄ¤═¼īWéāŻ¼┐╔ęį└ŁĄĮ╬──®Æ▀┤a╠Ē╝ėČYųŪąĪ┐═Ę■┼ČŻĪ

- LASERŽĄ┴ą╗Ņ¾w│╔Ž±ŽĄĮyų·┴”▄øŚŚ½J║’╠ę╗©ŪÓ╦žš{┐žÖCųŲĮŌ╬÷
- ¤ß³c╦┘▀f:Į³╝t═Ō╝{├ū╠Įßś║Ž│╔╝╝ąg’@ų°╠ß╔²ŲõĮ³╝t═Ō░l╣ŌÅŖČ╚┼cĘĆČ©ąį
- žō▌dBMSCį┤ąįĄ“═÷─ę┼▌Ą─└wŠSų¦╝▄═©▀^šTī¦Š▐╩╔╝Ü░¹śO╗»┤┘▀Mé¹┐┌ė·║Ž
- Š▀ėą░ļ╚ķ╠Ū░ąŽ“Ą─NIR-II╝{├ū╠ĮßśīŹ¼FįŁ╬╗Ė╬░®╩ųągŪąŠē╝░╔ŅČ╚Š½£╩│╔Ž±
- šō╬─ĮŌūxŻ║╣”─▄│¼┬Ģ│╔Ž±Ż©fUSŻ®ė├ė┌│¼įńŲ┌ūõųąļAČ╬▒O£yĄ─蹊┐░Ė└²
- ╬─½I╦┘▀fŻ║Ēö┐»CNS╔±ĮøŅIė“蹊┐ą┬▀Mš╣11į┬Ż©Ž┬Ż®
- └¹ė├1880-2080╝{├ū┤░┐┌▀MąąĖ▀ī”▒╚Č╚╗Ņ¾w¤╔╣Ō│╔Ž±Ą─ą┬ĘĮĘ©
- ╣®¾wŲĮ├µąįšTī¦Ęųūė┼żŪ·śŗĮ©D-A-Dą═AIEīŹ¼FĖ▀ą¦NIR-II¤╔╣Ō┼c╣Ō¤ßų╬»¤
- Iconeus┼cĘ©ć°ßtīW╬’└Ē蹊┐╦∙śs½@═╗ŲŲąįäōą┬¬ä
- ▓®·ś“vķWę½┴┴ŽÓ├└ć°īŹ“×äė╬’┐ŲīWģfĢ■AALAS╚½ć°Ģ■ūh
- ę½╦┘┐Ų╝╝┼c╚µ┌ĄŽ▀_│╔æ┬į║Žū„Ż¼╣▓═Ų╦Ä╬’čą░läōą┬
- Ū┌ŽĶIVScopeäė╬’╗Ņ¾w│╔Ž±ų·┴”ķ_░lmRNA-LNPŽĄĮy
- ▓®·ś“v┼cUCLA CRUMPĘųūėė░Ž±čąŠ┐╦∙įO┴ó╗Ņ¾wė░Ž±ųąą─
- ķ_─╗į┌╝┤Ż¼ICONEUSĄ╚╝┤īó┴┴ŽÓ├└ć°╔±Įø┐ŲīWīWĢ■─ĻĢ■
- Ū┌ŽĶīóöy╗Ņ¾w│╔Ž±Ą╚ģóš╣Ą┌╩«Ų▀ī├╚½ć°├Ōę▀īWīWąg┤¾Ģ■
- ICONEUS┼c─·ŽÓ╝s├└ć°╔±Įø┐ŲīWīWĢ■─ĻĢ■SfN2025



