
使用m6A-seq等揭示同義突變通過表觀轉(zhuǎn)錄調(diào)控機制決定生物性狀
近日,中國農(nóng)業(yè)科學院蔬菜花卉研究所楊學勇研究員、中國農(nóng)業(yè)科學院深圳農(nóng)業(yè)基因組所黃三文研究員和英國約翰英納斯中心丁一倞研究員團隊合作,以封面文章形式在頂刊《Cell》(細胞)上發(fā)表題為“Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation”的突破性前沿成果。研究聚焦黃瓜果實長度的馴化性狀,發(fā)現(xiàn)兩個緊密連鎖且存在上位互作的基因——FL1.1和FL1.2,分別由 YTH1 和 ACS2 基因定義,揭示了ACS2中的1287位C>T 同義突變(不改變氨基酸序列的DNA變異)通過介導m6A修飾變化和mRNA結(jié)構(gòu)構(gòu)象調(diào)控ACS2蛋白翻譯效率(TE),進而影響黃瓜果實長度的馴化機制,為理解同義突變的生物學功能提供了多細胞生物層面實例。

標題:Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation(同義突變的隱性上位效應通過表觀轉(zhuǎn)錄組調(diào)控賦予黃瓜馴化特性)
發(fā)表時間:2025年8月21日
發(fā)表期刊:Cell
影響因子:IF42.5/Q1
技術平臺:遺傳定位與基因克隆、基因編輯、m6A-seq、RNA結(jié)構(gòu)分析、免疫印跡與核糖體圖譜分析等
作者單位:楊學勇研究員、黃三文研究員和丁一倞研究員為本文共同通訊作者,中國農(nóng)業(yè)科學院蔬菜花卉研究所辛同旭博士、河南大學張震副教授、英國約翰英納斯中心張月瑩博士和中國農(nóng)業(yè)科學院蔬菜花卉研究所已畢業(yè)碩士生李旭彤為本文共同第一作者。
DOI:10.1016/j.cell.2025.06.007
長久以來,同義突變(Synonymous mutations)因不改變蛋白質(zhì)功能而被視為“沉默突變”。近期研究發(fā)現(xiàn)同義突變可能影響轉(zhuǎn)錄或轉(zhuǎn)錄后過程,但其對生物性狀的影響,尤其是在個體水平上的影響,仍有待深入研究。本研究鑒定出兩個緊密連鎖且存在上位互作的基因:YTH1(m6A reader蛋白編碼基因)和ACS2(ACC合成酶編碼基因),這兩個基因與黃瓜果實長度馴化高度相關。ACS2中的致病突變是1287位C>T同義突變。具體而言,在野生黃瓜(hardwickii)中,ACS2¹²⁸⁷C會導致腺苷殘基發(fā)生m6A修飾,并導致松散的RNA結(jié)構(gòu)構(gòu)象,從而提高ACS2蛋白表達水平,使果實變短;在栽培黃瓜(Xintaimici,新泰密刺)中,ACS2¹²⁸⁷T 會破壞m6A甲基化,形成緊密的RNA結(jié)構(gòu)構(gòu)象,導致蛋白表達量降低,果實變長。本研究為表觀轉(zhuǎn)錄組調(diào)控通過同義變異影響生物性狀提供了遺傳學證據(jù)。
研究亮點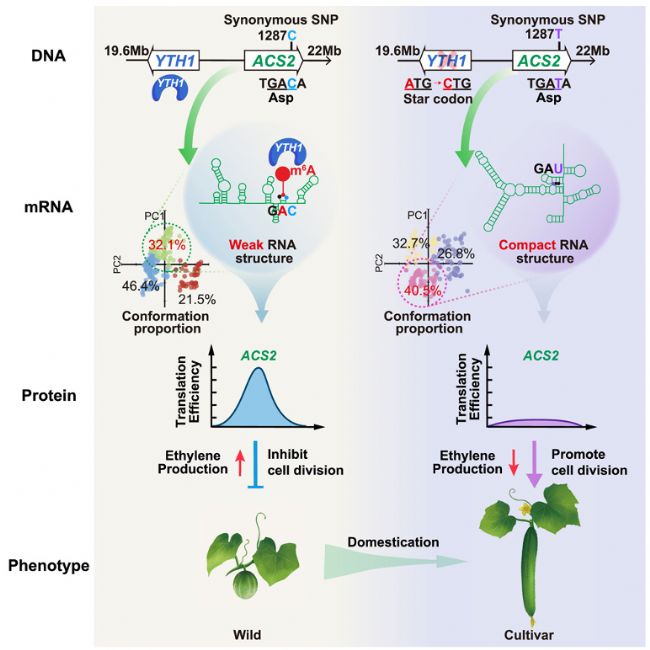
研究圖形摘要
易小結(jié)
本研究通過 m6A-seq 等技術揭示了黃瓜果實長度馴化的遺傳機制,證明了ACS2基因中的1287位C>T同義突變通過改變m6A修飾和mRNA結(jié)構(gòu)調(diào)控ACS2蛋白的翻譯效率,進而影響果實長度。
m6A-seq在本研究中不僅揭示了ACS2 mRNA上的m6A修飾位點及其在馴化過程中的變化,還進一步闡明了m6A修飾與RNA結(jié)構(gòu)、翻譯效率之間的復雜關系。這不僅為作物馴化研究提供了新的思路,也為易基因的表觀基因組學技術在農(nóng)業(yè)生物領域的應用提供了重要參考。易基因的表觀基因組技術,如m6A-seq、ChIP-seq和WGBS等,能夠全面解析基因表達調(diào)控網(wǎng)絡,助力研究人員深入理解生物性狀的遺傳基礎,為作物改良和新品種培育提供科學依據(jù)。
研究方法
(1)與果實長度馴化相關的兩個緊密連鎖的互作基因
研究人員發(fā)現(xiàn)黃瓜果實長度這一馴化性狀由位于1號染色體的FL1位點調(diào)控,進一步將該位點細分為FL1.1和FL1.2兩個緊密連鎖的基因位點。通過構(gòu)建回交導入系(ILs)和F₂群體分析,發(fā)現(xiàn)FL1.1和FL1.2基因位點存在上位互作,且FL1.2對FL1.1位點具有隱性上位性效應。當FL1.2為栽培等位基因時,無論FL1.1的基因型,果實始終是長瓜。這表明黃瓜果實長度的馴化由這兩個緊密連鎖且互作的數(shù)量性狀位點(QTL)共同決定。
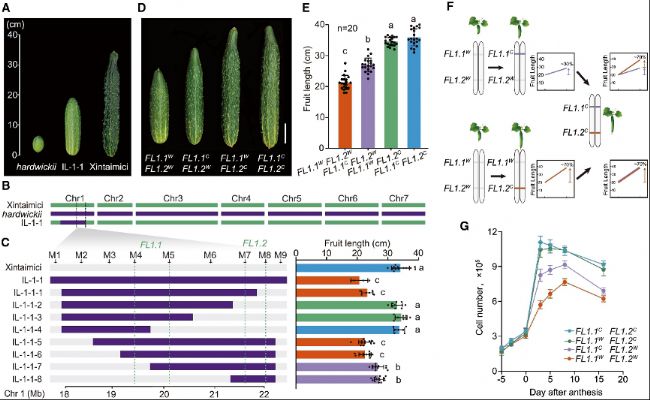
圖1:與果實長度馴化相關的兩個緊密連鎖、相互作用的基因
(A)野生黃瓜hardwickii(左側(cè))、栽培黃瓜Xintaimici(右側(cè))以及IL-1-1(中間)的代表性果實。
(B)代表性ILs和親本基因型。綠色和紫色分別代表Xintaimici和hardwickii的基因型。
(C)通過將IL-1-1漸滲片段細分為更小的片段(IL-1-1-1至IL-1-1-8),鑒定并定位兩個互作位點FL1.1和FL1.2。左側(cè)為選定重組體的基因型。灰色和紫色分別代表Xintaimici和hardwickii的基因型。綠色虛線定義了FL1.1和FL1.2的遺傳區(qū)間。右側(cè)為相應重組體植株的果實長度。
(D)四種基因型的代表性果實表型。W和C上標分別代表野生型和栽培型等位基因。
(E)與(D)相關的果實長度定量分析。
(F)模式圖顯示FL1.2C在遺傳上是隱性的,并對FL1.1具有上位性效應。紫色和紅色分別代表FL1.1C和FL1.2C的基因型。
(G)四種基因型的細胞數(shù)量分析。
(2)FL1.2由ACS2基因的同義突變定義
研究團隊將FL1.2定位到17.3kb區(qū)間,發(fā)現(xiàn)該區(qū)間僅含ACS2基因。ACS2編碼1-氨基環(huán)丙烷-1-羧酸合成酶,是乙烯合成的限速酶。基因組測序顯示,野生黃瓜和栽培黃瓜的ACS2存在多個變異,但只有1287位C>T同義突變顯著影響ACS2蛋白水平。通過雙熒光素酶實驗和轉(zhuǎn)基因?qū)嶒烌炞C,1287位C>T同義突變降低了ACS2蛋白水平,進而導致黃瓜果實變長,是FL1.2調(diào)控黃瓜果實長度馴化的因果突變。
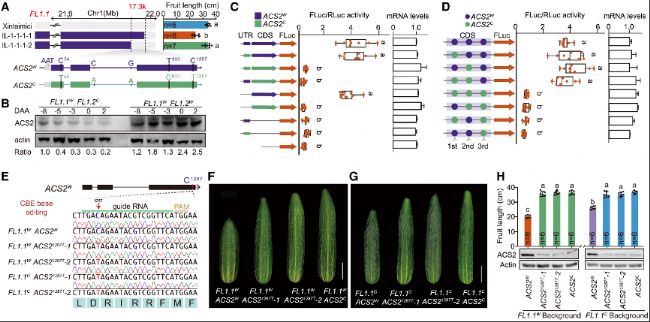
(B) 免疫印跡分析顯示早期果實發(fā)育過程中ACS2蛋白的水平。
(C) 雙熒光素酶報告基因分析對ACS2的5′UTR或CDS變異進行功能鑒定。左側(cè):示意圖顯示ACS2W和ACS2C的5′UTR和CDS變異的所有組合構(gòu)建。中間:左側(cè)顯示相應構(gòu)建的FLuc/RLuc活性。右側(cè):相應構(gòu)建的相對ACS2 mRNA表達水平。
(D) 使用雙熒光素酶報告基因分析法對三個同義SNP進行功能鑒定。左側(cè):示意圖顯示ACS2W和ACS2C中三個同義SNP的不同組合構(gòu)建。中間:左側(cè)顯示相應構(gòu)建的FLuc/RLuc活性。右側(cè):各種構(gòu)建的相對ACS2 mRNA表達水平。
(E) ACS2¹²⁸⁷C的堿基編輯結(jié)果。
(F-G) 帶有C1287到T轉(zhuǎn)換的轉(zhuǎn)基因植株果實長度。
(H) 與(F)和(G)相關的果實長度和ACS2蛋白水平的定量分析。
(3)FL1.1由YTH1基因的有害突變定義
FL1.1被定位到11kb區(qū)間,該區(qū)間僅含YTH1基因。YTH1編碼m6A reader蛋白。基因組測序發(fā)現(xiàn),栽培黃瓜的YTH1起始密碼子發(fā)生A→C突變,導致YTH結(jié)構(gòu)域缺失,轉(zhuǎn)錄水平顯著降低。通過基因敲除實驗驗證,YTH1是FL1.1的功能基因,且ACS2的栽培等位基因?qū)TH1存在隱性上位效應。這表明YTH1的有害突變通過影響m6A修飾的鑒定和調(diào)控,進而參與黃瓜果實長度的馴化過程。
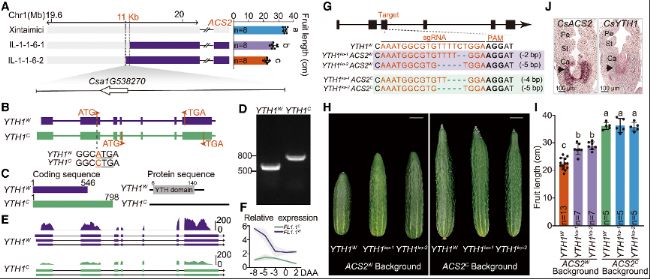
圖3:FL1.1由YTH1基因的有害突變定義
(A) FL1.1的精細定位區(qū)間包含YTH1基因。上:選定重組體的基因型和果實長度。灰色和紫色分別代表Xintaimici和hardwickii的基因型。
(B) YTH1基因結(jié)構(gòu)和變異。下方序列是起始密碼子周圍堿基。
(C) YTH1W和YTH1C的CDS和蛋白序列長度。數(shù)字表示CDS和YTH蛋白結(jié)構(gòu)域的起始和終止位置。
(D) PCR顯示YTH1W和YTH1C的CDS長度。
(E-F) 全長轉(zhuǎn)錄組(E)和RT-qPCR(F)顯示mRNA的剪切和表達情況。
(G) 使用單RNA(single-guide RNA, sgRNA)通過CRISPR-Cas9技術生成YTH1ko突變體。sgRNA靶點和PAM分別用紅色和加粗字體表示。缺失部分用藍色虛線表示。
(H-I) YTH1ko突變體的果實長度。
(J) 原位雜交實驗顯示發(fā)育中的子房心皮中ACS2和YTH1 mRNA的表達情況。Pe表示花瓣;St表示雄蕊;Ca表示心皮。
(4)1287位C>T同義突變調(diào)控ACS2 mRNA m6A甲基化和蛋白翻譯效率
通過m6A-seq分析發(fā)現(xiàn),ACS2 mRNA的第三外顯子存在顯著的m6A peaks,且1287位C>T同義突變顯著降低了ACS2 mRNA的m6A甲基化水平。進一步的免疫印跡和核糖體圖譜分析顯示,1287位C>T同義突變降低了ACS2蛋白水平和翻譯效率(TE),而YTH1的存在進一步增強了ACS2¹²⁸⁷C的TE。這表明1287位C>T同義突變通過影響m6A修飾和YTH1的結(jié)合,調(diào)控ACS2的蛋白翻譯效率,進而影響黃瓜果實長度。
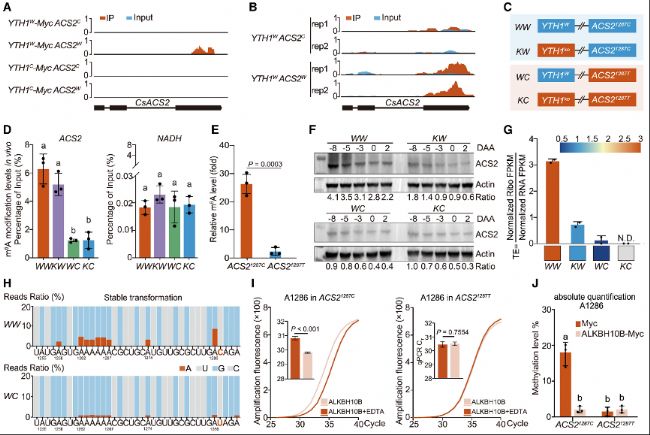
圖4:1287位C>T 同義突變調(diào)控ACS2 mRNA m6A甲基化和蛋白TE
(A) ACS2 mRNA的FA-CLIP富集peak覆蓋圖。
(B) ACS2 mRNA的m6A-seq富集peak覆蓋圖。
(C) 四個等基因系的培育,僅在YTH1基因和ACS2 1287同義突變上存在差異。
(D) m6A-RIP-qPCR檢測等基因系WW、KW、WC 和 KC中ACS2(左側(cè))和NADH(右側(cè)對照)的相對m6A水平。
(E) m6A-RIP-qPCR 檢測35S::ACS21287C 和35S::ACS21287T 的相對 m6A 水平。
(F) 四個等基因系中ACS2蛋白水平的免疫印跡分析。
(G) 核糖體分析法檢測WW、KW、WC 和KC中ACS2的相對翻譯效率(TE)。下:相應TE熱圖。
(H) 單堿基分辨率檢測 ACS2 mRNA 1287同義SNP附近的 m6A 修飾。
(I) ACS21287C (左)和 ACS21287T(右)中A1286的m6A修飾SELECT檢測結(jié)果。
(J) 絕對定量RT-PCR結(jié)果顯示ACS21287C 中A1286的m6A修飾水平。
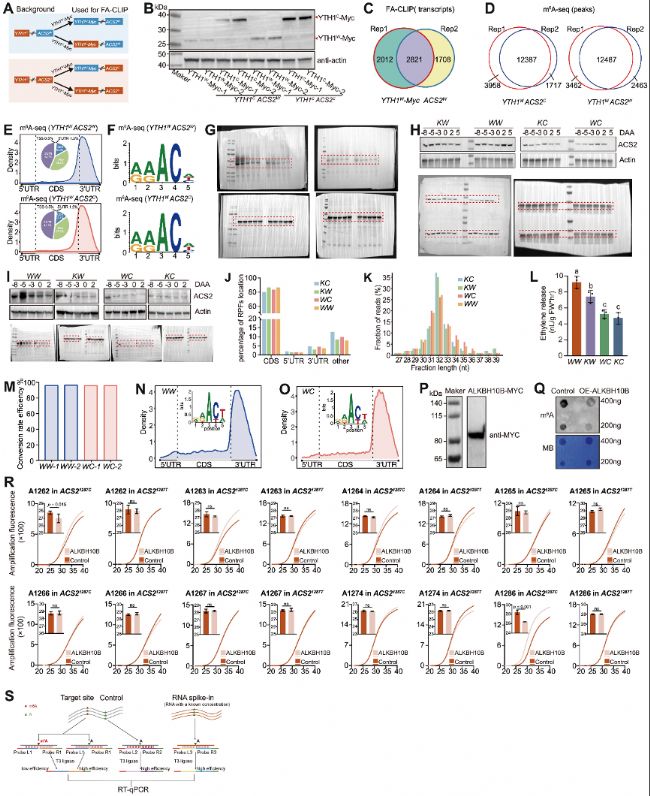
(B) 抗Myc抗體通過免疫印跡分析檢測轉(zhuǎn)基因材料中YTH1W-Myc 和YTH1C-Myc的蛋白表達。
(C) YTH1W-Myc W兩個生物學重復的FA-CLIP中YTH1結(jié)合轉(zhuǎn)錄本重疊。
(D) YTH1W-Myc ACS2W和YTH1W-Myc ACS2C兩個生物學重復m6A-seq中m6A甲基化peak重疊。
(E) m6A-seq鑒定的m6A位點在YTH1W-Myc ACS2W和YTH1W-Myc ACS2C轉(zhuǎn)錄本分布的meta基因分析。轉(zhuǎn)錄本分為三個區(qū)域:5′UTR、CDS和3′UTR。餅圖顯示在5′UTR、CDS和3′UTR區(qū)域分布的m6A位點比例(%)。
(F) 含有m6A的peak區(qū)域的保守序列motif。
(G–I) 與圖4F相關的三個重復的未裁剪免疫印跡圖像。
(J) 四種材料中核糖體足跡(RPFs)在CDS、5′UTR、3′UTR和其他區(qū)域的位置比例。
(K) RPFs長度比例分布。
(L) 四種基因型(WW、KW、WC和KC)中的乙烯產(chǎn)生量。
(M) 兩個獨立重復的eTAM-seq在轉(zhuǎn)錄組范圍內(nèi)的A→G轉(zhuǎn)換效率。
(N-O) 通過eTAM-seq鑒定的m6A位點在WW和WC轉(zhuǎn)錄本中的分布的meta基因分析。
(P) 抗MYC抗體的免疫印跡分析顯示ALKBH10B-MYC的表達。
(Q) 銅瓜同源的m6A去甲基化酶基因ALKBH10B過表達可以顯著降低黃瓜子葉中的全局m6A水平。
(R) 檢測ACS21287C和ACS21287T中1287同義突變周圍的全部腺苷m6A修飾的SELECT結(jié)果。
(5)1287位C>T同義突變通過m6A修飾和RNA結(jié)構(gòu)改變調(diào)控ACS2的TE
研究發(fā)現(xiàn)1287位C>T同義突變位于類似m6A保守motif DRACH的序列中,該突變破壞了m6A修飾位點,導致mRNA結(jié)構(gòu)從松散狀態(tài)變?yōu)榫o湊狀態(tài),降低了YTH1與ACS2 mRNA的結(jié)合能力,最終使ACS2的TE下降。而野生型1287C通過m6A修飾形成松散RNA結(jié)構(gòu),結(jié)合YTH1后進一步偏向最弱構(gòu)象,提高TE。這說明1287位C>T同義突變通過改變m6A修飾和RNA結(jié)構(gòu)構(gòu)象,精細調(diào)控ACS2翻譯效率,從而決定黃瓜果實長度的馴化性狀。
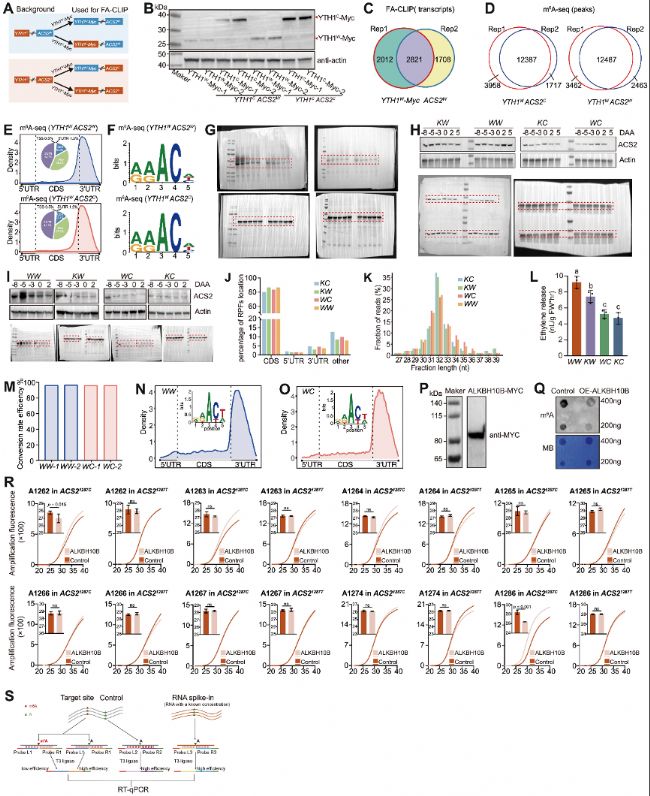
(B) FA-CLIP顯示了YTH1W與Myc、YTH1-Myc和ALKBH10B-Myc分別共表達時對ACS21287C和ACS21287T的結(jié)合能力。
(C) 雙熒光素酶報告基因分析顯示了與Myc、YTH1-Myc和ALKBH10B-Myc分別共表達時ACS21287C和ACS21287T的蛋白水平。
(D) ACS2等位基因的DMS反應性圖譜。
(E) DeltaSHAPE圖譜顯示了(D)中灰色陰影區(qū)域的DMS反應性差異,計算為每種條件與后續(xù)條件之間的差異:ACS21287T與ACS21287C(1287T與1287C)、與ALKBH10B共表達的ACS21287C與ACS21287C(ALKBH10B與1287C)、與YTH1共表達的ACS21287C與ACS21287C(YTH1與1287C)以及與ALKBH10B共表達的ACS21287C與ACS21287T(ALKBH10B與1287T)。
(F) ACS21287T和ACS21287C等位基因之間打開RNA結(jié)構(gòu)所需的能量差異。
(G) (C)中的TE與其對應的ΔG_unfold之間的皮爾遜相關性。
(H) CS21287T、ACS21287C以及分別與ALKBH10B或YTH1共表達的ACS21287C的代表性結(jié)構(gòu)模型。結(jié)構(gòu)通過主成分分析(PCA)可視化。
結(jié)論和啟示
本研究通過遺傳定位、基因克隆、基因編輯、m6A-seq分析等多種方法,揭示了黃瓜果實長度馴化性狀的遺傳調(diào)控機制,即ACS2基因中的1287位C>T同義突變通過改變m6A修飾和mRNA結(jié)構(gòu)構(gòu)象,調(diào)控ACS2蛋白的翻譯效率,進而影響黃瓜果實長度。另外,YTH1基因的有害突變通過影響m6A修飾的識別和調(diào)控,也參與了果實長度的馴化過程。這一研究結(jié)果不僅挑戰(zhàn)了同義突變無功能的傳統(tǒng)認知,拓展了m6A在同義突變的研究領域,還為作物遺傳改良提供了新的策略和思路。未來的研究可以進一步深入探索同義突變在其他生物性狀調(diào)控中的作用,以及m6A修飾與其他RNA修飾在基因表達調(diào)控中的相互作用機制,為生命科學研究和農(nóng)業(yè)生產(chǎn)實踐提供更多的理論支持和技術指導。
m6A-seq技術的關鍵作用
本研究通過m6A-seq分析,精確地定位ACS2 mRNA上的m6A修飾位點,并發(fā)現(xiàn)1287位C>T同義突變顯著降低了m6A修飾水平。這一發(fā)現(xiàn)為后續(xù)研究m6A修飾與RNA結(jié)構(gòu)、蛋白翻譯效率之間的關系提供了重要線索,進而揭示了m6A修飾在調(diào)控黃瓜果實長度馴化性狀中的關鍵作用。
在今后類似的研究中,m6A-seq技術有望得到更廣泛的應用。例如,在研究基因表達調(diào)控網(wǎng)絡時,m6A-seq可以與其他組學技術(如轉(zhuǎn)錄組測序、蛋白質(zhì)組測序等)相結(jié)合,全面解析m6A修飾在基因轉(zhuǎn)錄、剪接、翻譯等過程中的作用,以及其與其他調(diào)控因子之間的相互作用關系。此外,m6A-seq技術還可應用于研究發(fā)育生物學、疾病發(fā)生機制等領域,通過分析不同發(fā)育階段或疾病狀態(tài)下m6A修飾的動態(tài)變化,尋找潛在的調(diào)控靶點和生物標志物,為疾病的診斷和治療提供新的策略。
參考文獻:
Xin T, Zhang Z, Zhang Y, Li X, Wang S, Wang G, Li H, Wang B, Zhang M, Li W, Tian H, Zhang Z, Xiao YL, Tang W, He C, Ding Y, Huang S, Yang X. Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation. Cell. 2025 Jul 15. doi: 10.1016/j.cell.2025.07.006.
標題:Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation(同義突變的隱性上位效應通過表觀轉(zhuǎn)錄組調(diào)控賦予黃瓜馴化特性)
發(fā)表時間:2025年8月21日
發(fā)表期刊:Cell
影響因子:IF42.5/Q1
技術平臺:遺傳定位與基因克隆、基因編輯、m6A-seq、RNA結(jié)構(gòu)分析、免疫印跡與核糖體圖譜分析等
作者單位:楊學勇研究員、黃三文研究員和丁一倞研究員為本文共同通訊作者,中國農(nóng)業(yè)科學院蔬菜花卉研究所辛同旭博士、河南大學張震副教授、英國約翰英納斯中心張月瑩博士和中國農(nóng)業(yè)科學院蔬菜花卉研究所已畢業(yè)碩士生李旭彤為本文共同第一作者。
DOI:10.1016/j.cell.2025.06.007
長久以來,同義突變(Synonymous mutations)因不改變蛋白質(zhì)功能而被視為“沉默突變”。近期研究發(fā)現(xiàn)同義突變可能影響轉(zhuǎn)錄或轉(zhuǎn)錄后過程,但其對生物性狀的影響,尤其是在個體水平上的影響,仍有待深入研究。本研究鑒定出兩個緊密連鎖且存在上位互作的基因:YTH1(m6A reader蛋白編碼基因)和ACS2(ACC合成酶編碼基因),這兩個基因與黃瓜果實長度馴化高度相關。ACS2中的致病突變是1287位C>T同義突變。具體而言,在野生黃瓜(hardwickii)中,ACS2¹²⁸⁷C會導致腺苷殘基發(fā)生m6A修飾,并導致松散的RNA結(jié)構(gòu)構(gòu)象,從而提高ACS2蛋白表達水平,使果實變短;在栽培黃瓜(Xintaimici,新泰密刺)中,ACS2¹²⁸⁷T 會破壞m6A甲基化,形成緊密的RNA結(jié)構(gòu)構(gòu)象,導致蛋白表達量降低,果實變長。本研究為表觀轉(zhuǎn)錄組調(diào)控通過同義變異影響生物性狀提供了遺傳學證據(jù)。
研究亮點
- 兩個存在上位互作的基因YTH1和ACS2賦予黃瓜馴化特性。
- ACS2的1287位C>T同義突變是致病變異。
- 1287T破壞鄰近的m6A甲基化,并形成緊湊的ACS2 RNA結(jié)構(gòu)。
- m6A使RNA結(jié)構(gòu)構(gòu)象更弱,并增強翻譯效率。
研究圖形摘要
易小結(jié)
本研究通過 m6A-seq 等技術揭示了黃瓜果實長度馴化的遺傳機制,證明了ACS2基因中的1287位C>T同義突變通過改變m6A修飾和mRNA結(jié)構(gòu)調(diào)控ACS2蛋白的翻譯效率,進而影響果實長度。
m6A-seq在本研究中不僅揭示了ACS2 mRNA上的m6A修飾位點及其在馴化過程中的變化,還進一步闡明了m6A修飾與RNA結(jié)構(gòu)、翻譯效率之間的復雜關系。這不僅為作物馴化研究提供了新的思路,也為易基因的表觀基因組學技術在農(nóng)業(yè)生物領域的應用提供了重要參考。易基因的表觀基因組技術,如m6A-seq、ChIP-seq和WGBS等,能夠全面解析基因表達調(diào)控網(wǎng)絡,助力研究人員深入理解生物性狀的遺傳基礎,為作物改良和新品種培育提供科學依據(jù)。
研究方法
- 遺傳定位與基因克隆:利用野生黃瓜(hardwickii)和栽培黃瓜(Xintaimici)構(gòu)建回交導入系(introgression lines,ILs),通過精細定位將調(diào)控果實長度的FL1位點細分為FL1.1和FL1.2兩個緊密連鎖的基因位點,并進一步克隆出這兩個位點的關鍵基因YTH1和ACS2。
- 基因編輯:開發(fā)適用于瓜類作物的單堿基編輯工具,將ACS2野生等位基因的1287位的C編輯為T,直接驗證該同義突變的遺傳功能,證明其對黃瓜果實長度馴化的調(diào)控作用。
- m6A-seq和m6A-RIP-qPCR:通過m6A-seq分析ACS2 mRNA的m6A修飾水平,發(fā)現(xiàn)ACS2 mRNA的第三外顯子存在顯著的m6A peaks,且1287位C>T同義突變顯著降低ACS2 mRNA的m6A甲基化水平,揭示m6A修飾在調(diào)控ACS2蛋白翻譯效率中的關鍵作用。
- RNA結(jié)構(gòu)分析:采用單分子RNA結(jié)構(gòu)構(gòu)象多樣性分析(DAVINCI)等技術,分析ACS2 mRNA的結(jié)構(gòu)構(gòu)象多樣性,發(fā)現(xiàn)1287位C>T同義突變介導mRNA結(jié)構(gòu)從松散狀態(tài)變?yōu)榫o湊狀態(tài),進而影響YTH1與ACS2 mRNA的結(jié)合能力及ACS2翻譯效率。
- 免疫印跡與核糖體圖譜分析:通過免疫印跡檢測不同基因型植株中ACS2蛋白的水平,結(jié)合核糖體圖譜分析ACS2的翻譯效率(TE),驗證1287位C>T同義突變及YTH1對ACS2蛋白水平和TE的調(diào)控效應。
(1)與果實長度馴化相關的兩個緊密連鎖的互作基因
研究人員發(fā)現(xiàn)黃瓜果實長度這一馴化性狀由位于1號染色體的FL1位點調(diào)控,進一步將該位點細分為FL1.1和FL1.2兩個緊密連鎖的基因位點。通過構(gòu)建回交導入系(ILs)和F₂群體分析,發(fā)現(xiàn)FL1.1和FL1.2基因位點存在上位互作,且FL1.2對FL1.1位點具有隱性上位性效應。當FL1.2為栽培等位基因時,無論FL1.1的基因型,果實始終是長瓜。這表明黃瓜果實長度的馴化由這兩個緊密連鎖且互作的數(shù)量性狀位點(QTL)共同決定。
圖1:與果實長度馴化相關的兩個緊密連鎖、相互作用的基因
(B)代表性ILs和親本基因型。綠色和紫色分別代表Xintaimici和hardwickii的基因型。
(C)通過將IL-1-1漸滲片段細分為更小的片段(IL-1-1-1至IL-1-1-8),鑒定并定位兩個互作位點FL1.1和FL1.2。左側(cè)為選定重組體的基因型。灰色和紫色分別代表Xintaimici和hardwickii的基因型。綠色虛線定義了FL1.1和FL1.2的遺傳區(qū)間。右側(cè)為相應重組體植株的果實長度。
(D)四種基因型的代表性果實表型。W和C上標分別代表野生型和栽培型等位基因。
(E)與(D)相關的果實長度定量分析。
(F)模式圖顯示FL1.2C在遺傳上是隱性的,并對FL1.1具有上位性效應。紫色和紅色分別代表FL1.1C和FL1.2C的基因型。
(G)四種基因型的細胞數(shù)量分析。
(2)FL1.2由ACS2基因的同義突變定義
研究團隊將FL1.2定位到17.3kb區(qū)間,發(fā)現(xiàn)該區(qū)間僅含ACS2基因。ACS2編碼1-氨基環(huán)丙烷-1-羧酸合成酶,是乙烯合成的限速酶。基因組測序顯示,野生黃瓜和栽培黃瓜的ACS2存在多個變異,但只有1287位C>T同義突變顯著影響ACS2蛋白水平。通過雙熒光素酶實驗和轉(zhuǎn)基因?qū)嶒烌炞C,1287位C>T同義突變降低了ACS2蛋白水平,進而導致黃瓜果實變長,是FL1.2調(diào)控黃瓜果實長度馴化的因果突變。
圖2:FL1.2由ACS2基因的同義突變定義
(A) FL1.2的精細定位區(qū)間包含ACS2基因。上:選定重組體的基因型和果實長度。下:ACS2W和ACS2C的基因結(jié)構(gòu)和變異。方框、線條和灰色方框分別代表外顯子、內(nèi)含子和非翻譯區(qū)(UTRs)。(B) 免疫印跡分析顯示早期果實發(fā)育過程中ACS2蛋白的水平。
(C) 雙熒光素酶報告基因分析對ACS2的5′UTR或CDS變異進行功能鑒定。左側(cè):示意圖顯示ACS2W和ACS2C的5′UTR和CDS變異的所有組合構(gòu)建。中間:左側(cè)顯示相應構(gòu)建的FLuc/RLuc活性。右側(cè):相應構(gòu)建的相對ACS2 mRNA表達水平。
(D) 使用雙熒光素酶報告基因分析法對三個同義SNP進行功能鑒定。左側(cè):示意圖顯示ACS2W和ACS2C中三個同義SNP的不同組合構(gòu)建。中間:左側(cè)顯示相應構(gòu)建的FLuc/RLuc活性。右側(cè):各種構(gòu)建的相對ACS2 mRNA表達水平。
(E) ACS2¹²⁸⁷C的堿基編輯結(jié)果。
(F-G) 帶有C1287到T轉(zhuǎn)換的轉(zhuǎn)基因植株果實長度。
(H) 與(F)和(G)相關的果實長度和ACS2蛋白水平的定量分析。
(3)FL1.1由YTH1基因的有害突變定義
FL1.1被定位到11kb區(qū)間,該區(qū)間僅含YTH1基因。YTH1編碼m6A reader蛋白。基因組測序發(fā)現(xiàn),栽培黃瓜的YTH1起始密碼子發(fā)生A→C突變,導致YTH結(jié)構(gòu)域缺失,轉(zhuǎn)錄水平顯著降低。通過基因敲除實驗驗證,YTH1是FL1.1的功能基因,且ACS2的栽培等位基因?qū)TH1存在隱性上位效應。這表明YTH1的有害突變通過影響m6A修飾的鑒定和調(diào)控,進而參與黃瓜果實長度的馴化過程。
圖3:FL1.1由YTH1基因的有害突變定義
(B) YTH1基因結(jié)構(gòu)和變異。下方序列是起始密碼子周圍堿基。
(C) YTH1W和YTH1C的CDS和蛋白序列長度。數(shù)字表示CDS和YTH蛋白結(jié)構(gòu)域的起始和終止位置。
(D) PCR顯示YTH1W和YTH1C的CDS長度。
(E-F) 全長轉(zhuǎn)錄組(E)和RT-qPCR(F)顯示mRNA的剪切和表達情況。
(G) 使用單RNA(single-guide RNA, sgRNA)通過CRISPR-Cas9技術生成YTH1ko突變體。sgRNA靶點和PAM分別用紅色和加粗字體表示。缺失部分用藍色虛線表示。
(H-I) YTH1ko突變體的果實長度。
(J) 原位雜交實驗顯示發(fā)育中的子房心皮中ACS2和YTH1 mRNA的表達情況。Pe表示花瓣;St表示雄蕊;Ca表示心皮。
(4)1287位C>T同義突變調(diào)控ACS2 mRNA m6A甲基化和蛋白翻譯效率
通過m6A-seq分析發(fā)現(xiàn),ACS2 mRNA的第三外顯子存在顯著的m6A peaks,且1287位C>T同義突變顯著降低了ACS2 mRNA的m6A甲基化水平。進一步的免疫印跡和核糖體圖譜分析顯示,1287位C>T同義突變降低了ACS2蛋白水平和翻譯效率(TE),而YTH1的存在進一步增強了ACS2¹²⁸⁷C的TE。這表明1287位C>T同義突變通過影響m6A修飾和YTH1的結(jié)合,調(diào)控ACS2的蛋白翻譯效率,進而影響黃瓜果實長度。
圖4:1287位C>T 同義突變調(diào)控ACS2 mRNA m6A甲基化和蛋白TE
(B) ACS2 mRNA的m6A-seq富集peak覆蓋圖。
(C) 四個等基因系的培育,僅在YTH1基因和ACS2 1287同義突變上存在差異。
(D) m6A-RIP-qPCR檢測等基因系WW、KW、WC 和 KC中ACS2(左側(cè))和NADH(右側(cè)對照)的相對m6A水平。
(E) m6A-RIP-qPCR 檢測35S::ACS21287C 和35S::ACS21287T 的相對 m6A 水平。
(F) 四個等基因系中ACS2蛋白水平的免疫印跡分析。
(G) 核糖體分析法檢測WW、KW、WC 和KC中ACS2的相對翻譯效率(TE)。下:相應TE熱圖。
(H) 單堿基分辨率檢測 ACS2 mRNA 1287同義SNP附近的 m6A 修飾。
(I) ACS21287C (左)和 ACS21287T(右)中A1286的m6A修飾SELECT檢測結(jié)果。
(J) 絕對定量RT-PCR結(jié)果顯示ACS21287C 中A1286的m6A修飾水平。
圖S4:ACS2 mRNA中A1286的m6A甲基化檢測,與圖4相關
(A) FA-CLIP實驗材料制備。(B) 抗Myc抗體通過免疫印跡分析檢測轉(zhuǎn)基因材料中YTH1W-Myc 和YTH1C-Myc的蛋白表達。
(C) YTH1W-Myc W兩個生物學重復的FA-CLIP中YTH1結(jié)合轉(zhuǎn)錄本重疊。
(D) YTH1W-Myc ACS2W和YTH1W-Myc ACS2C兩個生物學重復m6A-seq中m6A甲基化peak重疊。
(E) m6A-seq鑒定的m6A位點在YTH1W-Myc ACS2W和YTH1W-Myc ACS2C轉(zhuǎn)錄本分布的meta基因分析。轉(zhuǎn)錄本分為三個區(qū)域:5′UTR、CDS和3′UTR。餅圖顯示在5′UTR、CDS和3′UTR區(qū)域分布的m6A位點比例(%)。
(F) 含有m6A的peak區(qū)域的保守序列motif。
(G–I) 與圖4F相關的三個重復的未裁剪免疫印跡圖像。
(J) 四種材料中核糖體足跡(RPFs)在CDS、5′UTR、3′UTR和其他區(qū)域的位置比例。
(K) RPFs長度比例分布。
(L) 四種基因型(WW、KW、WC和KC)中的乙烯產(chǎn)生量。
(M) 兩個獨立重復的eTAM-seq在轉(zhuǎn)錄組范圍內(nèi)的A→G轉(zhuǎn)換效率。
(N-O) 通過eTAM-seq鑒定的m6A位點在WW和WC轉(zhuǎn)錄本中的分布的meta基因分析。
(P) 抗MYC抗體的免疫印跡分析顯示ALKBH10B-MYC的表達。
(Q) 銅瓜同源的m6A去甲基化酶基因ALKBH10B過表達可以顯著降低黃瓜子葉中的全局m6A水平。
(R) 檢測ACS21287C和ACS21287T中1287同義突變周圍的全部腺苷m6A修飾的SELECT結(jié)果。
(5)1287位C>T同義突變通過m6A修飾和RNA結(jié)構(gòu)改變調(diào)控ACS2的TE
研究發(fā)現(xiàn)1287位C>T同義突變位于類似m6A保守motif DRACH的序列中,該突變破壞了m6A修飾位點,導致mRNA結(jié)構(gòu)從松散狀態(tài)變?yōu)榫o湊狀態(tài),降低了YTH1與ACS2 mRNA的結(jié)合能力,最終使ACS2的TE下降。而野生型1287C通過m6A修飾形成松散RNA結(jié)構(gòu),結(jié)合YTH1后進一步偏向最弱構(gòu)象,提高TE。這說明1287位C>T同義突變通過改變m6A修飾和RNA結(jié)構(gòu)構(gòu)象,精細調(diào)控ACS2翻譯效率,從而決定黃瓜果實長度的馴化性狀。
圖5:1287位C>T同義突變通過m6A修飾和RNA結(jié)構(gòu)改變調(diào)控ACS2的TE
(A) m6A-RIP-qPCR顯示了與Myc、YTH1-Myc和ALKBH10B-Myc分別共表達時ACS21287C和ACS21287T的m6A甲基化水平。(B) FA-CLIP顯示了YTH1W與Myc、YTH1-Myc和ALKBH10B-Myc分別共表達時對ACS21287C和ACS21287T的結(jié)合能力。
(C) 雙熒光素酶報告基因分析顯示了與Myc、YTH1-Myc和ALKBH10B-Myc分別共表達時ACS21287C和ACS21287T的蛋白水平。
(D) ACS2等位基因的DMS反應性圖譜。
(E) DeltaSHAPE圖譜顯示了(D)中灰色陰影區(qū)域的DMS反應性差異,計算為每種條件與后續(xù)條件之間的差異:ACS21287T與ACS21287C(1287T與1287C)、與ALKBH10B共表達的ACS21287C與ACS21287C(ALKBH10B與1287C)、與YTH1共表達的ACS21287C與ACS21287C(YTH1與1287C)以及與ALKBH10B共表達的ACS21287C與ACS21287T(ALKBH10B與1287T)。
(F) ACS21287T和ACS21287C等位基因之間打開RNA結(jié)構(gòu)所需的能量差異。
(G) (C)中的TE與其對應的ΔG_unfold之間的皮爾遜相關性。
(H) CS21287T、ACS21287C以及分別與ALKBH10B或YTH1共表達的ACS21287C的代表性結(jié)構(gòu)模型。結(jié)構(gòu)通過主成分分析(PCA)可視化。
結(jié)論和啟示
本研究通過遺傳定位、基因克隆、基因編輯、m6A-seq分析等多種方法,揭示了黃瓜果實長度馴化性狀的遺傳調(diào)控機制,即ACS2基因中的1287位C>T同義突變通過改變m6A修飾和mRNA結(jié)構(gòu)構(gòu)象,調(diào)控ACS2蛋白的翻譯效率,進而影響黃瓜果實長度。另外,YTH1基因的有害突變通過影響m6A修飾的識別和調(diào)控,也參與了果實長度的馴化過程。這一研究結(jié)果不僅挑戰(zhàn)了同義突變無功能的傳統(tǒng)認知,拓展了m6A在同義突變的研究領域,還為作物遺傳改良提供了新的策略和思路。未來的研究可以進一步深入探索同義突變在其他生物性狀調(diào)控中的作用,以及m6A修飾與其他RNA修飾在基因表達調(diào)控中的相互作用機制,為生命科學研究和農(nóng)業(yè)生產(chǎn)實踐提供更多的理論支持和技術指導。
m6A-seq技術的關鍵作用
本研究通過m6A-seq分析,精確地定位ACS2 mRNA上的m6A修飾位點,并發(fā)現(xiàn)1287位C>T同義突變顯著降低了m6A修飾水平。這一發(fā)現(xiàn)為后續(xù)研究m6A修飾與RNA結(jié)構(gòu)、蛋白翻譯效率之間的關系提供了重要線索,進而揭示了m6A修飾在調(diào)控黃瓜果實長度馴化性狀中的關鍵作用。
在今后類似的研究中,m6A-seq技術有望得到更廣泛的應用。例如,在研究基因表達調(diào)控網(wǎng)絡時,m6A-seq可以與其他組學技術(如轉(zhuǎn)錄組測序、蛋白質(zhì)組測序等)相結(jié)合,全面解析m6A修飾在基因轉(zhuǎn)錄、剪接、翻譯等過程中的作用,以及其與其他調(diào)控因子之間的相互作用關系。此外,m6A-seq技術還可應用于研究發(fā)育生物學、疾病發(fā)生機制等領域,通過分析不同發(fā)育階段或疾病狀態(tài)下m6A修飾的動態(tài)變化,尋找潛在的調(diào)控靶點和生物標志物,為疾病的診斷和治療提供新的策略。
參考文獻:
Xin T, Zhang Z, Zhang Y, Li X, Wang S, Wang G, Li H, Wang B, Zhang M, Li W, Tian H, Zhang Z, Xiao YL, Tang W, He C, Ding Y, Huang S, Yang X. Recessive epistasis of a synonymous mutation confers cucumber domestication through epitranscriptomic regulation. Cell. 2025 Jul 15. doi: 10.1016/j.cell.2025.07.006.
標簽:
RNA甲基化

- 單細胞擬時序分析技術的核心特征、原理、分析步驟及應用場景
- 現(xiàn)貨型新生抗原疫苗聯(lián)合ICI在晚期纖維板層型肝癌中展現(xiàn)治療潛力
- 解鎖細胞年齡的密碼:端粒檢測如何成為健康管理新視角?
- 多組學分析揭示DNA低甲基化定義人組織調(diào)節(jié)性T細胞的表觀遺傳適應性
- Polysome Profiling—解析翻譯調(diào)控機制的金標準
- Ribo-seq—檢測正在翻譯的RNA信息 連接轉(zhuǎn)錄組學與蛋白質(zhì)組學的橋梁
- Science:利用大規(guī)模并行核糖體分析發(fā)現(xiàn)泛病毒ORF
- 微量WGBS揭示DNA甲基化調(diào)控斑馬魚造血干細胞發(fā)育的表觀遺傳機制




Copyright(C) 1998-2025 生物器材網(wǎng) 電話:021-64166852;13621656896 E-mail:info@bio-equip.com