
mTOR 通路調(diào)控自噬的過程介紹

自噬一直是研究中的熱門話題,而 mTOR 通路是信號通路中的“明星”,兩者之間“剪不斷理還亂”,這個故事還要從“調(diào)控”說起……
mTOR 是細胞生長和代謝的主要調(diào)節(jié)分子,可促進合成代謝過程,如核糖體的生物發(fā)生 (Ribosome biogenesis) 以及蛋白質(zhì)、核苷酸、脂肪酸和脂質(zhì)的合成,并抑制分解代謝過程,如自噬。mTOR 信號的失調(diào)與許多人類疾病有關(guān),包括糖尿病、神經(jīng)退行性疾病和癌癥。
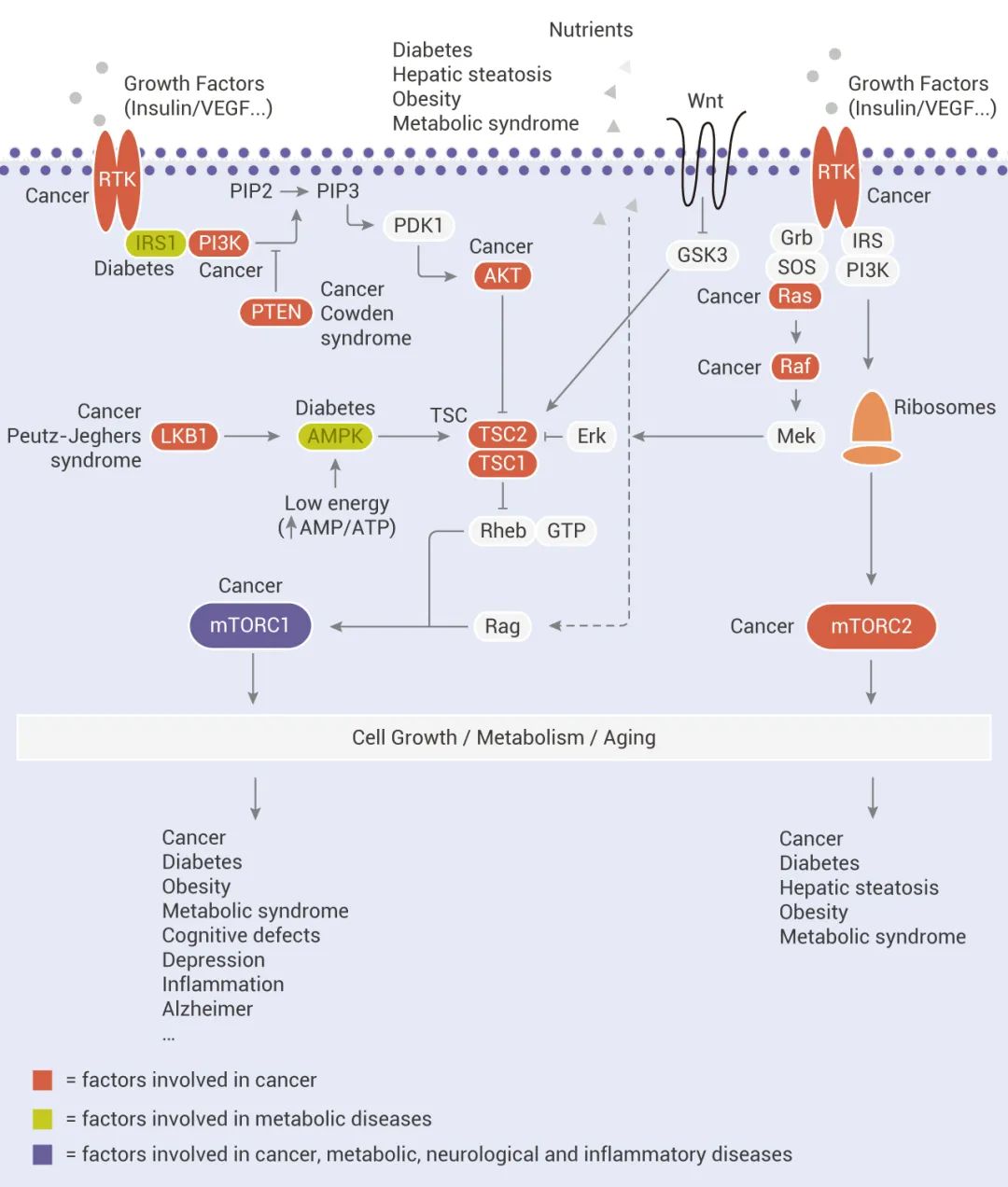
mTOR 是一種絲氨酸/蘇氨酸激酶,屬于 PI3K 相關(guān)激酶 (PIKK) 家族的成員,可與幾種蛋白質(zhì)相互作用形成兩個不同的復(fù)合物,分別稱為 mTOR 復(fù)合物 1 (mTORC1) 和 2 (mTORC2)。mTORC1 包含 mTOR、Raptor、PRAS40、mLST8 和 DEPTOR,對雷帕霉素 (Rapamycin) 敏感;mTORC2 包含 mTOR、Rictor、SIN1、Protor-1、mLST8 和 DEPTOR,對雷帕霉素急性治療不敏感。
mTOR 通過與 mTORC1 或 mTORC2 的關(guān)鍵組分如 Raptor、Rictor、LST8 和 SIN1 作用,催化多個靶標的磷酸化,如 S6K1、4E-BP1、Akt、PKC、 IGF-IR 和 ULK1,從而調(diào)節(jié)蛋白合成、營養(yǎng)代謝、生長因子信號傳導(dǎo)、細胞生長和遷移以及調(diào)節(jié)自噬等。
mTOR1/2 的活化
mTORC1 受多種刺激 (例如生長因子、營養(yǎng)物質(zhì)、能量和應(yīng)激信號) 以及基本信號傳導(dǎo)途徑 (例如 PI3K、MAPK 和 AMPK) 激活。
1、生長因子和激素 (例如胰島素) 通過 I 類 PI3K 及其下游效應(yīng)子 AKT 的激活來調(diào)節(jié) mTORC1 信號傳導(dǎo),從而逆轉(zhuǎn) TSC1/TSC2 復(fù)合物和 PRAS40 對 mTORC1 信號傳導(dǎo)的抑制作用;
2、營養(yǎng)物質(zhì)如氨基酸通過促進 RAS 相關(guān) GTP 結(jié)合蛋白 (RAG) 異二聚體向活性構(gòu)象的轉(zhuǎn)化來刺激 mTORC1;
3、細胞能量狀態(tài)如低 ATP 水平下,通過 AMPK 磷酸化 TSC2,mTORC1 活性受到抑制。除 PI3K 和 AMPK 途徑外,RAS/MAPK 信號也觸發(fā)了 mTORC1 信號的激活。此外,Hippo 途徑 (Hippo 途徑與靶向策略)和 Wnt 途徑 (Wnt 信號通路很復(fù)雜?) 也參與了對 mTORC1 和 mTORC2 的調(diào)控。
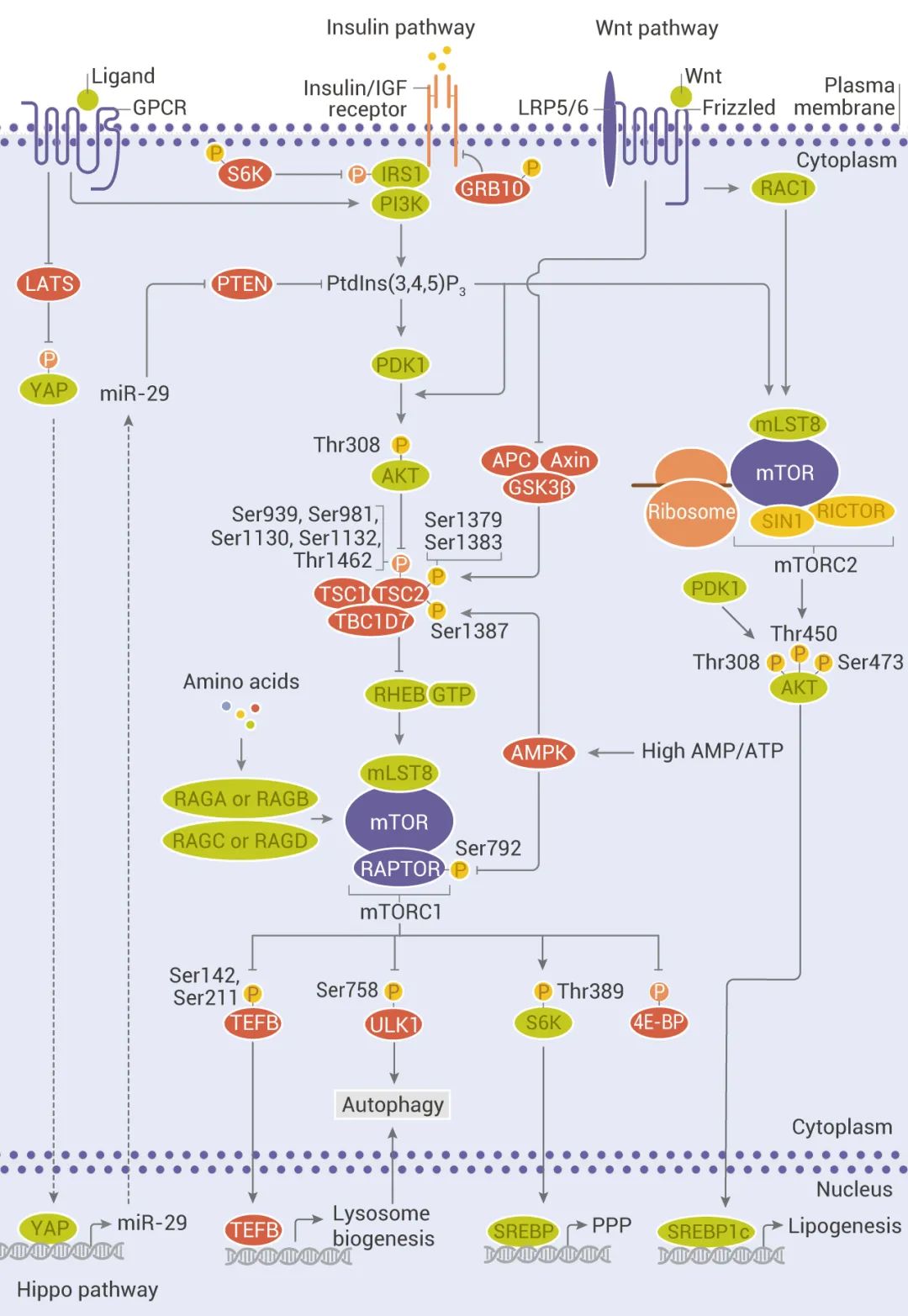
圖 2. mTOR 上游:經(jīng)典和非經(jīng)典輸入[1]
mTORC1 活化后,通過磷酸化下游效應(yīng)因子 S6K 和 4E-BP 促進合成代謝過程,例如蛋白質(zhì)、脂質(zhì)和核苷酸合成,同時,通過 ULK1 抑制分解代謝程序,從而導(dǎo)致自噬抑制。與 mTORC1 相比,對 mTORC2 的了解較少。如 mTORC2 可防止 AKT 泛素化和降解。mTORC2 還可激活 SREBP1c 促進成脂基因表達,以及刺激胰島素樣生長因子 2 (IGF2) 的產(chǎn)生。
mTOR 與自噬
自噬是一種溶酶體降解系統(tǒng),涉及從頭自噬形成。而自噬體的形成是由一系列稱為 Atg 或“自噬相關(guān)”蛋白的蛋白質(zhì)的協(xié)同作用驅(qū)動的。成熟的自噬體與溶酶體融合后,會被酸化,形成自溶酶體。溶酶體與自噬體的融合降解捕獲的蛋白質(zhì),脂質(zhì),碳水化合物,核酸和細胞器 (Hello~自噬)。
大量研究表明 mTORC1 是自噬的關(guān)鍵調(diào)節(jié)劑,調(diào)節(jié)自噬過程中不同步驟 (如成核,自噬體延伸,自噬體成熟和終止) 。因此,mTORC1 是調(diào)控自噬的極具潛力的靶標。
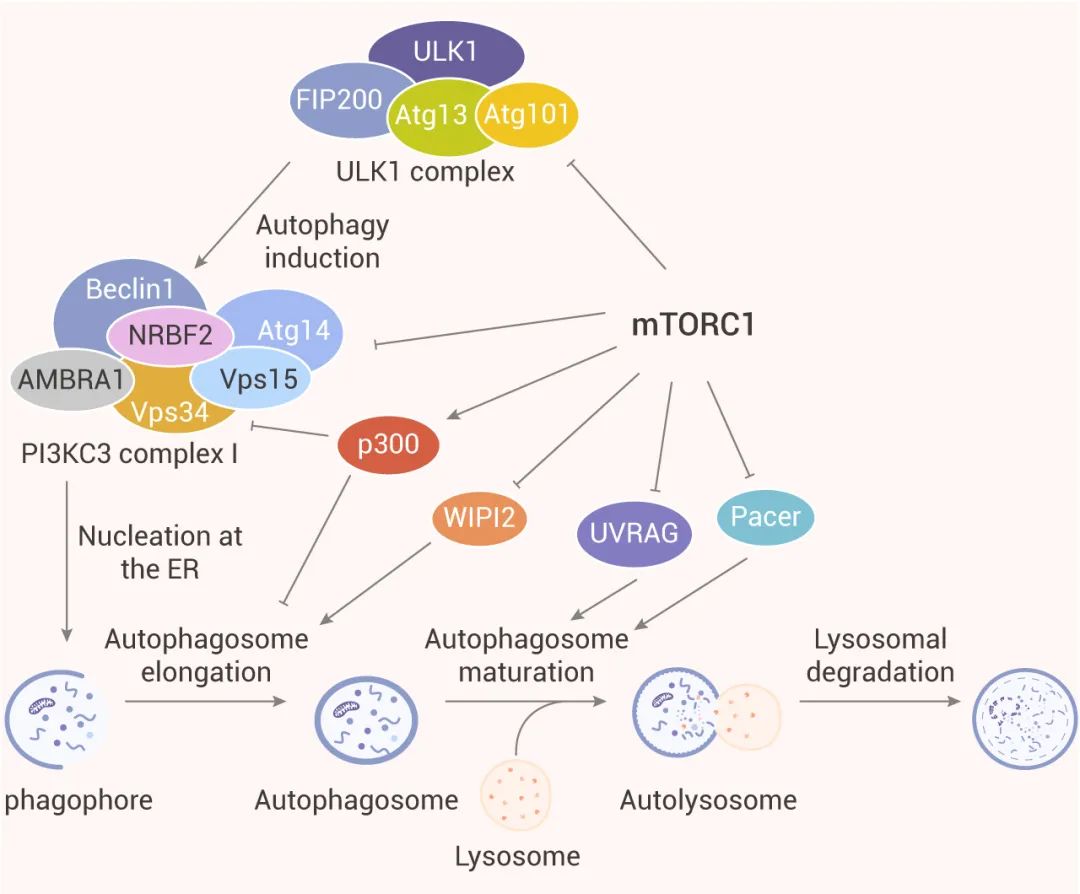
圖 3. mTORC1 對自噬各個步驟的調(diào)控[9]
mTORC1 如何調(diào)控自噬呢?
1、研究表明,mTORC1 通過磷酸化使形成的自噬調(diào)節(jié)復(fù)合物 (由 ULK1 和其互作蛋白 Atg13、FIP200、Atg101 等形成) 失活,從而影響自噬小體的生物發(fā)生。在營養(yǎng)豐富的條件下,mTORC1 通過介導(dǎo) ULK1 (Ser637 和 Ser757) 和 Atg13 (Ser258) 特定位點磷酸化,抑制 ULK1 復(fù)合物的自噬促進激酶 (Autophagy-promoting kinase) 活性。在饑餓和細胞應(yīng)激期間,mTORC1 活性被抑制,于是與 ULK1 分離。因此,ULK1 和 Atg13 的特定位點的磷酸化被解除。同時,ULK1 復(fù)合物通過 Thr180 處自磷酸化而變得活躍,并磷酸化 Atg13、FIP200、Atg101 和其他 Atg 蛋白。活躍的 ULK1 復(fù)合物隨后轉(zhuǎn)移到內(nèi)質(zhì)網(wǎng)的隔離膜上,自噬啟動。
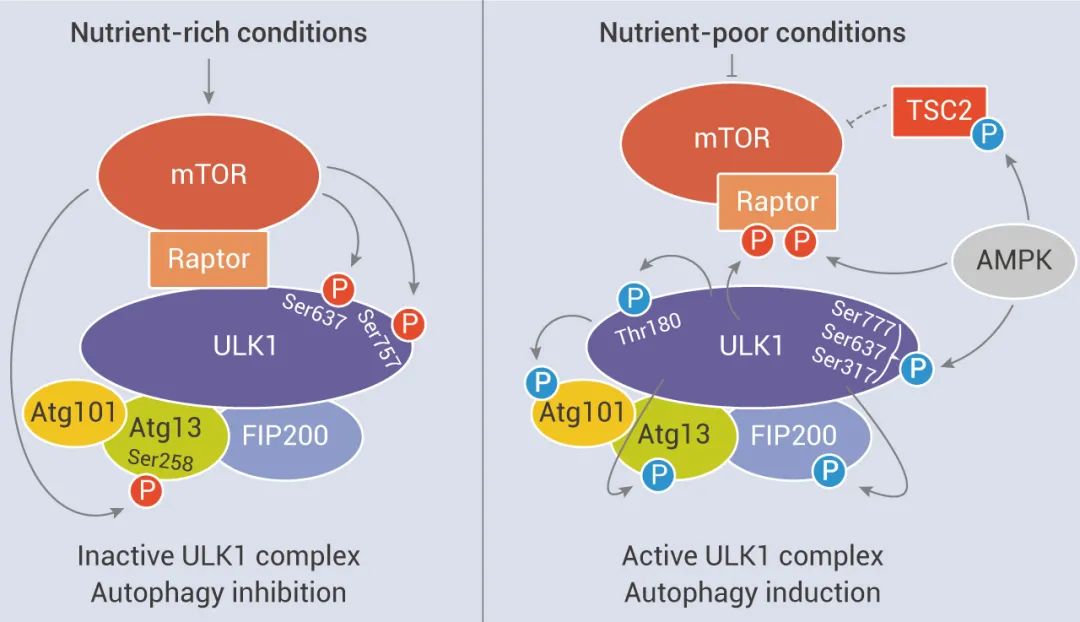
2. 另外,AMPK 也與自噬有關(guān)。在葡萄糖充足的情況下,活性 mTORC1 通過磷酸化 ULK1 特定位點 (Ser 757) 并破壞 ULK1 與 AMPK 之間的相互作用來阻止 ULK1 活化,抑制自噬啟動。在葡萄糖不足的情況下,AMPK 被激活,mTORC1 的磷酸化而被 AMPK 抑制,隨后 ULK1 可以與 AMPK 相互作用并被 AMPK 磷酸化,活化的 ULK1 啟動自噬。
3、自噬的成核步驟受到 PI3KC3 復(fù)合體 I 中 Atg14、AMBRA1 和 NRBF2 的磷酸化抑制。因此,mTORC1 可以通過磷酸化其成分 Atg14,AMBRA1 和 NRBF2 直接調(diào)節(jié) PI3KC3-CI 的活性。
4、研究表明,mTORC1 通過分別靶向 WIPI2 和 p300 乙酰轉(zhuǎn)移酶參與調(diào)節(jié)自噬體形成的延伸步驟,以及 LC3 與自噬體膜的結(jié)合。如通過抑制 p300 分子內(nèi)自抑制,可促進 LC3 的乙酰化,這阻礙了 LC3 的脂化。WIPI2 通過與 Atg16L 結(jié)合并將 Atg12-Atg5-Atg16L 復(fù)合物結(jié)合到吞噬細胞上來促進磷脂酰乙醇胺對 LC3 的脂化作用。
5、mTORC1 還可以通過調(diào)節(jié)溶酶體生物發(fā)生所需基因的轉(zhuǎn)錄來間接抑制自噬。如 TFEB 是溶酶體生物發(fā)生和自噬基因的主要轉(zhuǎn)錄調(diào)節(jié)因子。TFEB 可上調(diào)與自噬體形成、自噬體與溶酶體的融合相關(guān),以及溶酶體生物發(fā)生所需的一系列基因。此外,TFEB 的過表達可增加 UVRAG、WIPI、MAPLC3B、SQSTM1、VPS11、VPS19 和 ATG9B 的表達,它們參與了自噬的各個步驟。此外,mTORC2 也可通過激活 mTORC1 間接抑制自噬。
由于自噬與運動、代謝適應(yīng)和衰老等生理過程,以及神經(jīng)退行性疾病、傳染病、心血管疾病,癌癥等疾病過程有關(guān),但自噬誘導(dǎo)是利是弊,取決于疾病的類型或階段。例如,當(dāng)營養(yǎng)有限時,自噬可以通過向腫瘤細胞提供營養(yǎng)來促進腫瘤發(fā)生和發(fā)展。因此,自噬的抑制可能使癌細胞對代謝應(yīng)激條件敏感,從而導(dǎo)致細胞死亡。另外,mTORC1 作為自噬的主要調(diào)節(jié)劑,參與自噬過程,是極具有潛力靶標,實際上,mTORC1 的失調(diào)已經(jīng)牽涉到與自噬缺陷相關(guān)的疾病中,并且已經(jīng)有 mTOR 抑制劑在臨床試驗中或獲批用于治療這些疾病。
總之,mTOR 促進合成代謝并抑制自噬誘導(dǎo),而用 mTOR 抑制劑調(diào)節(jié)自噬為多種疾病提供了新的治療策略。
|
具有誘導(dǎo)自噬作用的 mTOR 抑制劑 |
| 有效且特異性的 mTOR 抑制劑,作用于 HEK293 細胞,抑制 mTOR,IC50 為 0.1 nM;與 FKBP12 結(jié)合且抑制 mTORC1;還是一種自噬 (autophagy) 激活劑,免疫抑制劑。 |
| mTOR 抑制劑,IC50 值為 1.76 μM;能激活自噬 (autophagy),在動物模型中防止心臟功能惡化。 |
| ATP 競爭性的 mTOR 抑制劑,IC50 為 5 nM;能抑制 mTORC1 和 mTORC2;在體外能誘導(dǎo)自噬 (autophagy) 激活。 |
| 抑制肝臟中的線粒體呼吸鏈,導(dǎo)致 AMPK 活化,增強胰島素敏感性,可用于 2 型糖尿病的研究;可以透過血腦屏障,誘導(dǎo)自噬 (autophagy)。 |
| Cell Counting Kit-8,簡稱CCK-8試劑盒或CCK8試劑盒,是一種基于WST-8而廣泛應(yīng)用于細胞活性和細胞毒性檢測的快速、高靈敏度試劑盒。 |
MCE 的所有產(chǎn)品僅用作科學(xué)研究或藥證申報,我們不為任何個人用途提供產(chǎn)品和服務(wù)
參考文獻
↓ 下滑查看更多文獻
1. Shimobayashi M, et al. Making new contacts: the mTOR network in metabolism and signalling crosstalk. Nat Rev Mol Cell Biol. 2014; 15(3): 155-162.
2. Dazert E, et al. mTOR signaling in disease. Curr Opin Cell Biol. 2011; 23(6): 744-755.
3. Hua H, et al. Targeting mTOR for cancer therapy. J Hematol Oncol. 2019; 12(1): 71. Published 2019 Jul 5.
4. Mossmann D, et al. mTOR signalling and cellular metabolism are mutual determinants in cancer. Nat Rev Cancer. 2018; 18(12): 744-757.
5. Alayev A, et al. mTOR signaling for biological control and cancer. J Cell Physiol. 2013; 228(8): 1658-1664.
6. Kim J, et al. AMPK and mTOR regulate autophagy through direct phosphorylation of ULK1. Nat Cell Biol. 2011; 13(2): 132-141.
7. Wang H, et al. The Upstream Pathway of mTOR-Mediated Autophagy in Liver Diseases. Cells. 2019; 8(12): 1597.
8. Kim YC, et al. mTOR: a pharmacologic target for autophagy regulation. J Clin Invest. 2015; 125(1): 25-32.
9. Akpedje S. Dossou, et al. The Emerging Roles of mTORC1 in Macromanaging Autophagy. Cancers (Basel). 2019 Oct; 11(10): 1422.
10. Paquette M, et al. mTOR Pathways in Cancer and Autophagy. Cancers (Basel). 2018; 10(1): 18.
11. Laplante M, et al. mTOR signaling in growth control and disease. Cell. 2012; 149(2): 274-293.

- 受擠壓的細胞記住自身形狀以高效遷移的機制科普
- 全新細胞死亡方式Mitoxyperilysis的機制及其在疾病治療中的價值
- 實驗性自身免疫性腦脊髓炎(EAE)造模誘導(dǎo)方法及核心試劑的優(yōu)勢和作用
- 細胞因子IL-10 、IL-12、干擾素及腫瘤壞死因子 (TNF) 家族介紹及應(yīng)用
- 受體酪氨酸激酶 (RTKs)結(jié)構(gòu)組成、激活機制及其在抗癌研究中的應(yīng)用
- OCT包埋劑的作用原理、使用方法及應(yīng)用
- PBS緩沖液的功能、作用機制及在生命科學(xué)研究中的多場景應(yīng)用
- Dexamethasone地塞米松(DEX)在構(gòu)建動物疾病模型中的應(yīng)用



