
具有更強免疫原性的mRNA疫苗介紹
1 mRNA簡介
上個世紀九十年代,在一系列體外轉錄技術被突破的背景下,mRNA可以作為治療藥物的概念正式被提出來。隨后,由于結構簡單、安全性好,以及可以誘導機體免疫反應的特點,mRNA技術逐漸被開發為疫苗產品平臺。
理論上,mRNA疫苗具有眾多傳統疫苗不具備的優點。首先,跟傳統疫苗相比較mRNA具有更強的免疫原性。mRNA可以表達特定序列的靶蛋白誘導特異性免疫反應,同時作為核酸類的免疫原,mRNA還可以誘導人體的天然免疫反應(見下文),使其擁有“自我佐劑”(self-adjuvant)的特點,兩方面的作用,使得疫苗具有更強的免疫原性;第二,在安全性方面,mRNA疫苗的遞送不需要特定的質粒載體,不進入細胞核內部,只需要在細胞質內瞬時表達抗原蛋白,不存在整合到人體基因組上的風險。第三,mRNA疫苗具有生產優勢。在成本上mRNA疫苗的GMP生產成本是傳統疫苗的五到十分之一。另外,由于化學結構相似,生產和純化與抗原本身無關,同一條mRNA疫苗的生產線很容易被改造成符合EMA或FDA標準的不同抗原疫苗的生產線,因此,在應對突發傳染病時,mRNA疫苗很容易在短時間內完成研發和生產。
雖然在理論上mRNA疫苗有著突出的優點,但是目前研發最快的產品還處于臨床試驗階段。阻礙mRNA疫苗技術發展的zui大障礙是其對先天性免疫的激活,這種免疫反應對mRNA疫苗來說是一把雙刃劍。一方面mRNA通過免疫激活來誘導機體的免疫保護,另一方面過度的激活先天性免疫可以阻止mRNA的翻譯,并降解mRNA(見下文)。從成藥性的角度來看,一款合格的mRNA疫苗產品要求其mRNA結構足夠穩定,能夠在靶細胞中遞送和表達并誘發預期的適應性免疫反應。
2 mRNA誘導的先天性免疫反應
先天性免疫是人體免疫系統的第一道防線,可以通過模式識別受體(Pattern Recognition Receptor,PRR)識別入侵抗原的病原相關模式分子(Pathogen-Associated Molecular Patterns,PAMP),然后通過一系列復雜的細胞內部級聯反應進行免疫應答。mRNA疫苗作為外源的核酸物質,進入體內后會被一系列位于細胞表面、內質網以及細胞質中的模式識別受體識別,并激發機體的先天性免疫應答,正如上文所述,這種免疫應答是一把“雙刃劍”。
研究發現,mRNA疫苗誘導的先天性免疫反應可以促進人體樹突狀細胞(Dendritic cell,DC)的成熟。成熟的DC進一步向人體的免疫系統遞呈mRNA表達的靶抗原,誘導機體產生特異的T細胞和B細胞免疫,對人體產生預期的免疫保護效應。從這個角度看,mRNA誘導的先天性免疫反應是有利的。
mRNA疫苗作為一種單鏈RNA其本身也是一種PRR,其單鏈結構和代謝產物可以激活細胞內的Toll樣受體7(TLR-7)和Toll樣受體8(TLR-8),并最終激活I型干擾素介導的先天性免疫反應,最終阻止mRNA表達并降解mRNA。另外,在體外進行mRNA合成的過程中會產生雙鏈RNA(double-strand RNA,dsRNA)。dsRNA也是一種常見的的核酸類PAMP,污染了dsRNA的mRNA產品可以上調并激活蛋白激酶R和寡聚腺苷酸合成酶,隨后通過I型干擾素介導的免疫反應阻止mRNA的翻譯及降解mRNA。從這個角度來看,mRNA誘導的先天性免疫反應對產品本身有損害作用。這種有害的免疫反應可以通過調節mRNA的核酸序列,在純化階段采用先進工藝去除dsRNA等手段避免。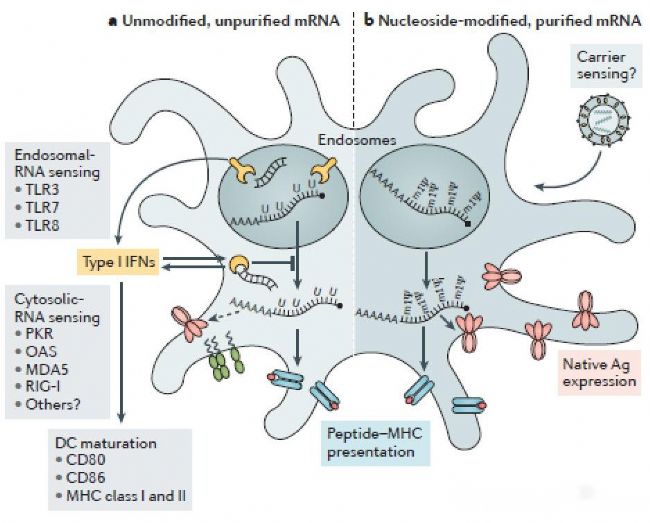
3 mRNA的結構及優化策略
3.1 mRNA加帽
真核細胞mRNA的5'端有一個帽子結構,它是在mRNA轉錄后,由甲基化的鳥苷酸(m7G)經焦磷酸化與mRNA的5'端核苷酸相連。5'端帽子結構在mRNA許多生物學功能中起到重要的作用,這包括:mRNA的剪切、穩定、轉運和核糖體的募集等。由于5'端帽子結構的重要生物學功能,在mRNA疫苗研發過程中需要優化5'端帽子結構,并利用合適的工藝對mRNA原料進行“加帽”。
mRNA疫苗產品的加帽可以在mRNA的體外轉錄過程中通過摻入“帽子序列”實現,這種加帽技術會將加帽序列反向加在mRNA上,這會導致約1/3的加帽失敗。通過摻入特殊的帽子結構——“anti-reverse”帽子結構(ARCA)可以降低這種反向加帽失誤的出現。另外,還可以先合成不帶帽子結構的mRNA,再利用痘病毒的“加帽復合物”對mRNA進行加帽,這也是一種mRNA的加帽手段。
3.2 非翻譯區序列(UTRs)修飾
在mRNA的編碼區兩端各存在一段非翻譯區(UTRs),在5'端的被稱為5'端UTR,在3'端的被稱為3'端UTR。
非翻譯區序列不是密碼子,不能翻譯成氨基酸,但是可以通過RNA結合蛋白控制mRNA產品的降解和轉錄效率。因此,每一種mRNA疫苗都需要考慮如何設計產品的UTR組件,進而提高在目的細胞中mRNA疫苗的穩定性和轉錄效率。大量的研究發現非洲爪蟾和人源的α-球蛋白/β-球蛋白的UTR序列非常穩定,而且能夠增加異源的mRNA的表達效率,因此這兩個蛋白的UTR原件通常被設計為mRNA疫苗的UTR組件。當然,也有其它蛋白的UTRs被研究用于mRNA疫苗的組件,在此不再贅述。
3.3 Poly(A)
大多數真核生物的mRNA 的3'端都有由100~200個腺苷酸聚合形成的Poly(A)尾巴。mRNA的Poly(A)尾巴可以和5'端帽子結構共同合作,增強mRNA結構的穩定性。Poly(A)尾巴首先和大量的Poly(A)結合蛋白(PABP)結合,然后募集真核生物起始因子4G(eIF4G),增加Poly(A)跟5'端帽子結構的親和力,最終mRNA的Poly(A)尾巴和5'端帽子結構首尾相接,形成環裝mRNA。環裝mRNA結構有利于核糖體的募集,其特殊的結構也能保護mRNA不被降解。
在mRNA體外合成過程中,Poly(A)尾巴可以通過質粒DNA模板轉錄獲得,也可以先合成mRNA再通過Poly(A)聚合酶加上不同單位數量的尾巴。前者能夠在mRNA產品中加入特定數目的Poly(A),而后者加入的單位數不確定。
Poly(A)尾巴的單位數目對于其生物學作用起到了至關重要的作用。在哺乳動物細胞內,活躍表達的mRNA包含的Poly(A)序列的長度為100-250個。有研究發現,表達效率越高的mRNA,其Poly(A)尾巴越短。但是當Poly(A)單位長度小于12時,mRNA結構就不再穩定,容易被降解。在設計mRNA疫苗產品的序列時,Poly(A)的長度也是非常重要的優化指標。
3.4 開放閱讀框
開放閱讀框(Open Reading Frame, ORF)包涵了mRNA的全部編碼序列,負責進入人體后表達mRNA運載的抗原靶蛋白。
密碼子水平的修飾,利用常用的密碼子,選用tRNA較多的密碼子;使各個密碼子的比例更接近靶物種或者細胞的比例;
GC含量的修飾,避免復雜二級結構形成;二級結構對mRNA的延伸和轉錄非常重要,高穩定的二級結構和發夾結構需要避免。這樣的二級結構可以阻止核糖體的進入、掃描以及容易被先天性免疫識別為PAMP。
4 mRNA疫苗分類
mRNA疫苗包括兩類:自我擴增型(self-amplifying or replicon)以及非復制型(non-replicating or synthetic)的mRNA疫苗。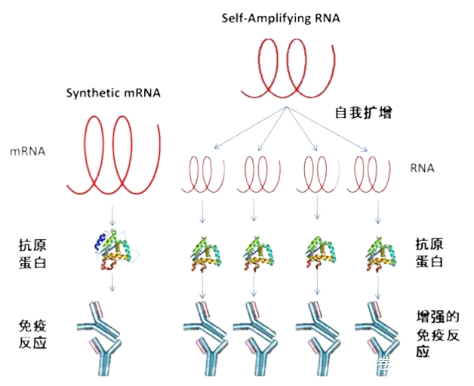
自我擴增型mRNA疫苗的設計通常是基于甲病毒屬(alphavirus)的病毒基因組。甲病毒屬為感染昆蟲的RNA病毒,用編碼抗原蛋白的mRNA代替甲病毒基因組上的甲病毒結構蛋白RNA,保留了甲病毒基因組自我復制的能力,又不能合成甲病毒蛋白。這類改造后的甲病毒基因組進入體內后可以自我復制產生編碼抗原蛋白的mRNA,很少的量就可以引起較強的免疫反應。
非復制型RNA疫苗則是在體外轉錄好的一段編碼抗原蛋白的完整mRNA,包括5ʹ和3ʹ的未翻譯區(UTRs),poly A tail用于穩定mRNA和促進轉錄;同時mRNA還有多種堿基修飾來提高mRNA的穩定性,zui后利用納米脂質體等遞送技術將mRNA遞送至細胞內,從而翻譯抗原蛋白,引發免疫反應,但是這種合成的mRNA在人體內無法自我復制。
參考文獻
DOI:10.1016/j.molmed.2019.10.002
DOI:10.4155/fsoa-2017-0151
DOI:10.1517/14712598.2015.1057563
DOI:org/10.1002/stem.2402
DOI:org/10.4161/rna.22269
DOI:org/10.3389/fimmu.2019.00594
DOI:10.1038/nrd.2017.243
DOI:10.1007/978-1-4939-6481-9
DOI:10.1186/s12967-016-1111-6
DOI:10.1038/s41541-020-0159-8
上個世紀九十年代,在一系列體外轉錄技術被突破的背景下,mRNA可以作為治療藥物的概念正式被提出來。隨后,由于結構簡單、安全性好,以及可以誘導機體免疫反應的特點,mRNA技術逐漸被開發為疫苗產品平臺。
理論上,mRNA疫苗具有眾多傳統疫苗不具備的優點。首先,跟傳統疫苗相比較mRNA具有更強的免疫原性。mRNA可以表達特定序列的靶蛋白誘導特異性免疫反應,同時作為核酸類的免疫原,mRNA還可以誘導人體的天然免疫反應(見下文),使其擁有“自我佐劑”(self-adjuvant)的特點,兩方面的作用,使得疫苗具有更強的免疫原性;第二,在安全性方面,mRNA疫苗的遞送不需要特定的質粒載體,不進入細胞核內部,只需要在細胞質內瞬時表達抗原蛋白,不存在整合到人體基因組上的風險。第三,mRNA疫苗具有生產優勢。在成本上mRNA疫苗的GMP生產成本是傳統疫苗的五到十分之一。另外,由于化學結構相似,生產和純化與抗原本身無關,同一條mRNA疫苗的生產線很容易被改造成符合EMA或FDA標準的不同抗原疫苗的生產線,因此,在應對突發傳染病時,mRNA疫苗很容易在短時間內完成研發和生產。
雖然在理論上mRNA疫苗有著突出的優點,但是目前研發最快的產品還處于臨床試驗階段。阻礙mRNA疫苗技術發展的zui大障礙是其對先天性免疫的激活,這種免疫反應對mRNA疫苗來說是一把雙刃劍。一方面mRNA通過免疫激活來誘導機體的免疫保護,另一方面過度的激活先天性免疫可以阻止mRNA的翻譯,并降解mRNA(見下文)。從成藥性的角度來看,一款合格的mRNA疫苗產品要求其mRNA結構足夠穩定,能夠在靶細胞中遞送和表達并誘發預期的適應性免疫反應。
2 mRNA誘導的先天性免疫反應
先天性免疫是人體免疫系統的第一道防線,可以通過模式識別受體(Pattern Recognition Receptor,PRR)識別入侵抗原的病原相關模式分子(Pathogen-Associated Molecular Patterns,PAMP),然后通過一系列復雜的細胞內部級聯反應進行免疫應答。mRNA疫苗作為外源的核酸物質,進入體內后會被一系列位于細胞表面、內質網以及細胞質中的模式識別受體識別,并激發機體的先天性免疫應答,正如上文所述,這種免疫應答是一把“雙刃劍”。
研究發現,mRNA疫苗誘導的先天性免疫反應可以促進人體樹突狀細胞(Dendritic cell,DC)的成熟。成熟的DC進一步向人體的免疫系統遞呈mRNA表達的靶抗原,誘導機體產生特異的T細胞和B細胞免疫,對人體產生預期的免疫保護效應。從這個角度看,mRNA誘導的先天性免疫反應是有利的。
mRNA疫苗作為一種單鏈RNA其本身也是一種PRR,其單鏈結構和代謝產物可以激活細胞內的Toll樣受體7(TLR-7)和Toll樣受體8(TLR-8),并最終激活I型干擾素介導的先天性免疫反應,最終阻止mRNA表達并降解mRNA。另外,在體外進行mRNA合成的過程中會產生雙鏈RNA(double-strand RNA,dsRNA)。dsRNA也是一種常見的的核酸類PAMP,污染了dsRNA的mRNA產品可以上調并激活蛋白激酶R和寡聚腺苷酸合成酶,隨后通過I型干擾素介導的免疫反應阻止mRNA的翻譯及降解mRNA。從這個角度來看,mRNA誘導的先天性免疫反應對產品本身有損害作用。這種有害的免疫反應可以通過調節mRNA的核酸序列,在純化階段采用先進工藝去除dsRNA等手段避免。
mRNA介導的先天性免疫反應
3 mRNA的結構及優化策略
3.1 mRNA加帽
真核細胞mRNA的5'端有一個帽子結構,它是在mRNA轉錄后,由甲基化的鳥苷酸(m7G)經焦磷酸化與mRNA的5'端核苷酸相連。5'端帽子結構在mRNA許多生物學功能中起到重要的作用,這包括:mRNA的剪切、穩定、轉運和核糖體的募集等。由于5'端帽子結構的重要生物學功能,在mRNA疫苗研發過程中需要優化5'端帽子結構,并利用合適的工藝對mRNA原料進行“加帽”。
mRNA疫苗產品的加帽可以在mRNA的體外轉錄過程中通過摻入“帽子序列”實現,這種加帽技術會將加帽序列反向加在mRNA上,這會導致約1/3的加帽失敗。通過摻入特殊的帽子結構——“anti-reverse”帽子結構(ARCA)可以降低這種反向加帽失誤的出現。另外,還可以先合成不帶帽子結構的mRNA,再利用痘病毒的“加帽復合物”對mRNA進行加帽,這也是一種mRNA的加帽手段。
3.2 非翻譯區序列(UTRs)修飾
在mRNA的編碼區兩端各存在一段非翻譯區(UTRs),在5'端的被稱為5'端UTR,在3'端的被稱為3'端UTR。
非翻譯區序列不是密碼子,不能翻譯成氨基酸,但是可以通過RNA結合蛋白控制mRNA產品的降解和轉錄效率。因此,每一種mRNA疫苗都需要考慮如何設計產品的UTR組件,進而提高在目的細胞中mRNA疫苗的穩定性和轉錄效率。大量的研究發現非洲爪蟾和人源的α-球蛋白/β-球蛋白的UTR序列非常穩定,而且能夠增加異源的mRNA的表達效率,因此這兩個蛋白的UTR原件通常被設計為mRNA疫苗的UTR組件。當然,也有其它蛋白的UTRs被研究用于mRNA疫苗的組件,在此不再贅述。
3.3 Poly(A)
大多數真核生物的mRNA 的3'端都有由100~200個腺苷酸聚合形成的Poly(A)尾巴。mRNA的Poly(A)尾巴可以和5'端帽子結構共同合作,增強mRNA結構的穩定性。Poly(A)尾巴首先和大量的Poly(A)結合蛋白(PABP)結合,然后募集真核生物起始因子4G(eIF4G),增加Poly(A)跟5'端帽子結構的親和力,最終mRNA的Poly(A)尾巴和5'端帽子結構首尾相接,形成環裝mRNA。環裝mRNA結構有利于核糖體的募集,其特殊的結構也能保護mRNA不被降解。
在mRNA體外合成過程中,Poly(A)尾巴可以通過質粒DNA模板轉錄獲得,也可以先合成mRNA再通過Poly(A)聚合酶加上不同單位數量的尾巴。前者能夠在mRNA產品中加入特定數目的Poly(A),而后者加入的單位數不確定。
Poly(A)尾巴的單位數目對于其生物學作用起到了至關重要的作用。在哺乳動物細胞內,活躍表達的mRNA包含的Poly(A)序列的長度為100-250個。有研究發現,表達效率越高的mRNA,其Poly(A)尾巴越短。但是當Poly(A)單位長度小于12時,mRNA結構就不再穩定,容易被降解。在設計mRNA疫苗產品的序列時,Poly(A)的長度也是非常重要的優化指標。
3.4 開放閱讀框
開放閱讀框(Open Reading Frame, ORF)包涵了mRNA的全部編碼序列,負責進入人體后表達mRNA運載的抗原靶蛋白。
密碼子水平的修飾,利用常用的密碼子,選用tRNA較多的密碼子;使各個密碼子的比例更接近靶物種或者細胞的比例;
GC含量的修飾,避免復雜二級結構形成;二級結構對mRNA的延伸和轉錄非常重要,高穩定的二級結構和發夾結構需要避免。這樣的二級結構可以阻止核糖體的進入、掃描以及容易被先天性免疫識別為PAMP。
4 mRNA疫苗分類
mRNA疫苗包括兩類:自我擴增型(self-amplifying or replicon)以及非復制型(non-replicating or synthetic)的mRNA疫苗。
mRNA疫苗的兩種類型
自我擴增型mRNA疫苗的設計通常是基于甲病毒屬(alphavirus)的病毒基因組。甲病毒屬為感染昆蟲的RNA病毒,用編碼抗原蛋白的mRNA代替甲病毒基因組上的甲病毒結構蛋白RNA,保留了甲病毒基因組自我復制的能力,又不能合成甲病毒蛋白。這類改造后的甲病毒基因組進入體內后可以自我復制產生編碼抗原蛋白的mRNA,很少的量就可以引起較強的免疫反應。
非復制型RNA疫苗則是在體外轉錄好的一段編碼抗原蛋白的完整mRNA,包括5ʹ和3ʹ的未翻譯區(UTRs),poly A tail用于穩定mRNA和促進轉錄;同時mRNA還有多種堿基修飾來提高mRNA的穩定性,zui后利用納米脂質體等遞送技術將mRNA遞送至細胞內,從而翻譯抗原蛋白,引發免疫反應,但是這種合成的mRNA在人體內無法自我復制。
參考文獻
DOI:10.1016/j.molmed.2019.10.002
DOI:10.4155/fsoa-2017-0151
DOI:10.1517/14712598.2015.1057563
DOI:org/10.1002/stem.2402
DOI:org/10.4161/rna.22269
DOI:org/10.3389/fimmu.2019.00594
DOI:10.1038/nrd.2017.243
DOI:10.1007/978-1-4939-6481-9
DOI:10.1186/s12967-016-1111-6
DOI:10.1038/s41541-020-0159-8





Copyright(C) 1998-2025 生物器材網 電話:021-64166852;13621656896 E-mail:info@bio-equip.com