
白細(xì)胞介素-2(IL-2)的功能、信號(hào)調(diào)控機(jī)制及對(duì)免疫穩(wěn)態(tài)調(diào)控的雙面性
在免疫系統(tǒng)的復(fù)雜調(diào)控網(wǎng)絡(luò)中,白細(xì)胞介素-2(IL-2)無(wú)疑是核心樞紐之一。自1976年被發(fā)現(xiàn)并命名為“T細(xì)胞生長(zhǎng)因子”以來(lái),IL-2憑借其既能驅(qū)動(dòng)免疫攻擊、又能維持免疫耐受的“雙面性”,成為連接基礎(chǔ)免疫學(xué)與臨床治療的關(guān)鍵分子。它通過(guò)精密的受體分級(jí)系統(tǒng)、多層次的信號(hào)通路調(diào)控,在抗腫瘤、抗感染與自身免疫平衡中發(fā)揮著不可替代的作用,同時(shí)也為免疫治療的創(chuàng)新提供了豐富的靶點(diǎn)與思路。
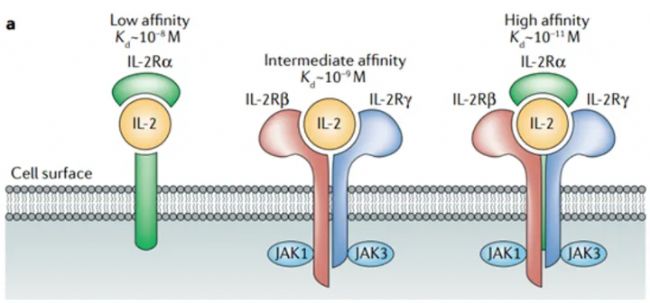
一、 分子基礎(chǔ):受體結(jié)構(gòu)決定功能特異性
IL-2的功能實(shí)現(xiàn),首先依賴于其與細(xì)胞表面受體(IL-2R)的特異性結(jié)合,而IL-2R的“分級(jí)設(shè)計(jì)”是其功能分化的核心基礎(chǔ)。IL-2R由α鏈(CD25)、β鏈(CD122)和γ鏈(CD132,又稱γc)三種亞基組成,三者以不同組合形成低、中、高三種親和力受體,分別對(duì)應(yīng)不同免疫細(xì)胞的功能需求(見下表)。
| 受體類型 | 亞基組成 | 親和力(Kd) | 主要表達(dá)細(xì)胞 | 核心功能 |
| 低親和力受體 | 僅 α 鏈(CD25) | 10⁻⁷~10⁻⁶ M | 部分活化免疫細(xì)胞 | “捕獲” IL-2,形成局部細(xì)胞因子庫(kù) |
| 中親和力受體 | β 鏈 +γ 鏈(CD122+CD132) | 10⁻⁹ M | 靜息效應(yīng) T 細(xì)胞(Teff)、NK 細(xì)胞、記憶 T 細(xì)胞(Tmem) | 響應(yīng)中高濃度 IL-2,啟動(dòng)免疫激活 |
| 高親和力受體 | α 鏈 +β 鏈 +γ 鏈(CD25+CD122+CD132) | 10⁻¹¹ M | 調(diào)節(jié)性 T 細(xì)胞(Treg)、活化 T 細(xì)胞 | 響應(yīng)低濃度 IL-2,維持 Treg 存活與功能 |
這種分級(jí)結(jié)構(gòu)的精妙之處在于:高親和力受體僅在Treg表面穩(wěn)定表達(dá),即使體內(nèi)IL-2濃度極低(10-100 U/mL),也能高效捕獲信號(hào),確保免疫耐受的基礎(chǔ);而中親和力受體主要分布于效應(yīng)免疫細(xì)胞,需較高濃度IL-2(>100 U/mL)才能激活,避免免疫反應(yīng)過(guò)度激活。例如,靜息狀態(tài)下的CD8+ T細(xì)胞僅表達(dá)中親和力受體,當(dāng)感染或腫瘤發(fā)生時(shí),局部IL-2濃度升高,才會(huì)啟動(dòng)其增殖與細(xì)胞毒性功能,實(shí)現(xiàn)“按需激活”。
二、 信號(hào)調(diào)控:三條核心通路構(gòu)建免疫平衡網(wǎng)絡(luò)
IL-2與受體結(jié)合后,通過(guò)激活JAK-STAT、PI3K-AKT-mTOR、MAPK-ERK三條核心信號(hào)通路,實(shí)現(xiàn)對(duì)T細(xì)胞命運(yùn)的精準(zhǔn)調(diào)控,三條通路既獨(dú)立作用又相互協(xié)同,共同維持免疫激活與耐受的動(dòng)態(tài)平衡。
(一)JAK-STAT通路:免疫耐受的“守護(hù)者”
IL-2結(jié)合受體后,首先誘導(dǎo)β鏈與γ鏈的細(xì)胞質(zhì)結(jié)構(gòu)域聚合,激活關(guān)聯(lián)的JAK1(結(jié)合β鏈)與JAK3(結(jié)合γ鏈)激酶。磷酸化的JAK進(jìn)而使STAT5蛋白磷酸化,形成二聚體后入核,調(diào)控關(guān)鍵基因表達(dá):一方面,促進(jìn)Treg特異性轉(zhuǎn)錄因子Foxp3的穩(wěn)定表達(dá),增強(qiáng)Treg的免疫抑制功能;另一方面,上調(diào)抗凋亡蛋白Bcl-2,延長(zhǎng)Treg的存活時(shí)間。研究證實(shí),STAT5缺陷小鼠會(huì)因Treg發(fā)育障礙引發(fā)嚴(yán)重自身免疫病,直接證明該通路是免疫耐受的核心保障。
(二)PI3K-AKT-mTOR通路:效應(yīng)T細(xì)胞的“動(dòng)力源”
該通路主要負(fù)責(zé)為效應(yīng)T細(xì)胞的活化提供代謝支持。IL-2激活PI3K后,通過(guò)下游AKT激酶激活mTORC1復(fù)合物,啟動(dòng)糖酵解代謝重編程——原本依賴氧化磷酸化的靜息T細(xì)胞,會(huì)快速切換為糖酵解為主的代謝模式,為細(xì)胞增殖、顆粒酶B與IFN-γ等細(xì)胞毒性分子的合成提供能量。同時(shí),AKT還會(huì)抑制Foxo轉(zhuǎn)錄因子,阻斷其向Treg分化的潛能,確保效應(yīng)T細(xì)胞“專注”于免疫攻擊。例如,高劑量IL-2治療黑色素瘤時(shí),正是通過(guò)強(qiáng)力激活該通路,使腫瘤浸潤(rùn)Teff大量擴(kuò)增,提升殺傷效率。
(三)MAPK-ERK通路:免疫激活的“放大器”
ERK通路的核心作用是形成正反饋循環(huán),增強(qiáng)IL-2的免疫激活效應(yīng)。IL-2誘導(dǎo)ERK磷酸化后,會(huì)促進(jìn)c-Fos與c-Jun蛋白形成AP-1轉(zhuǎn)錄復(fù)合體,上調(diào)IL-2Rα(CD25)的表達(dá)——這使得活化T細(xì)胞能更快組裝高親和力受體,進(jìn)一步增強(qiáng)對(duì)IL-2的響應(yīng)。此外,ERK還能通過(guò)磷酸化表觀調(diào)控因子EZH2,抑制Polycomb復(fù)合體的組蛋白甲基轉(zhuǎn)移酶活性,使效應(yīng)基因(如IFN-γ、顆粒酶B)的染色質(zhì)處于開放狀態(tài),為快速表達(dá)做好準(zhǔn)備。
三、 功能特性:濃度梯度決定“免疫雙面性”
IL-2的“雙面性”本質(zhì)是濃度依賴性功能選擇:不同濃度的IL-2會(huì)優(yōu)先激活不同免疫細(xì)胞亞群,實(shí)現(xiàn)“精準(zhǔn)調(diào)控”,這一特性也成為其臨床應(yīng)用的核心依據(jù)。
(一)高濃度IL-2:抗腫瘤與抗感染的“利器”
當(dāng)體內(nèi)IL-2濃度超過(guò)100 U/mL時(shí)(如感染急性期或高劑量IL-2治療),會(huì)優(yōu)先結(jié)合效應(yīng)T細(xì)胞與NK細(xì)胞的中親和力受體,啟動(dòng)強(qiáng)力免疫攻擊:CD8+ T細(xì)胞大量增殖,分泌IFN-γ與顆粒酶B,增強(qiáng)對(duì)腫瘤細(xì)胞或感染細(xì)胞的殺傷;NK細(xì)胞的細(xì)胞毒性活性提升,直接清除異常細(xì)胞。臨床中,高劑量IL-2(60萬(wàn)IU/kg)已獲批用于治療轉(zhuǎn)移性黑色素瘤與腎細(xì)胞癌,客觀緩解率達(dá)16%,部分患者可獲得長(zhǎng)期緩解。但高濃度IL-2也會(huì)引發(fā)嚴(yán)重副作用,如血管滲漏綜合征(VLS)——過(guò)量激活的免疫細(xì)胞會(huì)釋放大量炎癥因子,破壞血管內(nèi)皮屏障,導(dǎo)致組織水腫與低血壓,需在嚴(yán)密監(jiān)護(hù)下使用。
(二)低濃度IL-2:自身免疫病的“調(diào)節(jié)劑”
當(dāng)IL-2濃度處于10-100 U/mL時(shí),會(huì)優(yōu)先被Treg的高親和力受體捕獲,選擇性增強(qiáng)免疫耐受。例如,在系統(tǒng)性紅斑狼瘡(SLE)患者中,低劑量IL-2(1-1.5萬(wàn)IU/d)治療可使外周血Treg比例從3%提升至8%,顯著降低疾病活動(dòng)度;在1型糖尿病患者中,低劑量IL-2能減少自身反應(yīng)性T細(xì)胞對(duì)胰島β細(xì)胞的攻擊,延緩病情進(jìn)展。這種“低劑量靶向Treg”的策略,避免了對(duì)效應(yīng)免疫細(xì)胞的過(guò)度抑制,成為自身免疫病治療的新方向。
(三)脈沖式IL-2:免疫記憶的“維持者”
生理狀態(tài)下,IL-2的分泌呈周期性脈沖模式,這種模式能有效維持記憶T細(xì)胞的存活。當(dāng)IL-2以脈沖形式刺激時(shí),會(huì)與IL-7、IL-15協(xié)同作用,激活記憶T細(xì)胞的代謝重編程——既不引發(fā)過(guò)度增殖,又能維持其對(duì)病原體的“記憶”。例如,在慢性病毒感染(如HIV)中,脈沖式IL-2治療可提升記憶T細(xì)胞的數(shù)量與功能,增強(qiáng)對(duì)病毒的長(zhǎng)期控制。
四、 IL-2因子檢測(cè)服務(wù)哪里有?
IL-2作為免疫系統(tǒng)的“雙面因子”,其分子機(jī)制的解析不僅深化了我們對(duì)免疫穩(wěn)態(tài)的理解,也為臨床治療提供了多維度的靶點(diǎn)。從傳統(tǒng)高劑量治療腫瘤、低劑量調(diào)控自身免疫,到工程化變體與靶向遞送技術(shù)的創(chuàng)新,IL-2的應(yīng)用正從“廣譜調(diào)控”向“精準(zhǔn)靶向”邁進(jìn)。 未來(lái),隨著對(duì)IL-2信號(hào)網(wǎng)絡(luò)與免疫細(xì)胞互作機(jī)制的進(jìn)一步探索,有望開發(fā)出更高效、更安全的IL-2療法:例如,結(jié)合免疫檢查點(diǎn)抑制劑(如PD-1抗體),增強(qiáng)效應(yīng)T細(xì)胞的腫瘤浸潤(rùn);或通過(guò)時(shí)序調(diào)控IL-2濃度,實(shí)現(xiàn)“先激活效應(yīng)細(xì)胞、后抑制Treg”的動(dòng)態(tài)治療。這些創(chuàng)新將推動(dòng)IL-2在腫瘤、自身免疫病、感染性疾病等領(lǐng)域的應(yīng)用,為免疫治療的發(fā)展注入新動(dòng)力。
LabEx多款現(xiàn)貨Panel支持IL-2因子檢測(cè)服務(wù),歡迎咨詢~
| Panel | 樂(lè)備實(shí)官網(wǎng)貨號(hào) | 技術(shù)平臺(tái) | 檢測(cè)指標(biāo) |
| 人炎癥10因子Panel | LXMH10-1 | MSD | IFN-γ,IL-1β,IL-2,IL-4,IL-6,IL-8,IL-10,IL-12p70,IL-13,TNF-α |
| 人炎癥10因子Panel | LXLBH10-1 | Luminex | IL-1 β/IL-1F2,IL-2,IL-4,IL-6 ,IL-8/CXCL8,IL-10,IL-12 p70,IL-13,TNF-α,IFN-γ |
| 小鼠炎癥10因子Panel | LXMM10-1 | MSD | IFN-γ,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-10,IL-12p70,KC/GRO,TNF-α |
| 小鼠炎癥10因子Panel | LXLBM10-1 | Luminex | IL-1 β/IL-1F2,IL-2,IL-4,IL-5,IL-6 ,IL-10,IL-12p70,CXCL1/GRO/α/KC/CINC-1,IFN-γ,TNF-α |
| 大鼠炎癥10因子Panel | LXLBR10-1 | Luminex | IL-1 β/IL-1F2,IL-2,IL-4,IL-5,IL-6 ,IL-10,IL-12p70,CXCL1/GRO/α/KC/CINC-1,IFN-γ,TNF-α |
| 小鼠細(xì)胞因子-23因子Panel | LXLBM23-1 | Luminex | Eotaxin/CCL11,G-CSF,GM-CSF,IFN-γ,IL-10,IL-12(p40),IL-12(p70),IL-13,IL-17A,IL-1α,IL-1β,IL-2,IL-3,IL-4,IL-5,IL-6,IL-9,GRO-α (Gro-a/KC/CXCL1),MCP-1/CCL2,MIP-1α/CCL3,MIP-1β,RANTES,TNF-α |
| 大鼠細(xì)胞因子-23因子Panel | LXLBR23-1 | Luminex | G-CSF,GM-CSF,GRO/KC,IFN-γ,IL-1α,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-7,IL-10,IL-12 (p70),IL-13,IL-17A,IL-18,M-CSF,MCP-1,MIP-1α,MIP-3α,RANTES,TNF-α,VEGF |
| 人細(xì)胞因子-27因子Panel | LXLBH27-1 | Luminex | G-CSF,GM-CSF,IFN-γ,IL-10,IL-12(p70),IL-13,IL-17A,IL-1β,IL-2,IL-4,IL-5,IL-6,IL-7,IL-8/CXCL8,MCP-1/CCL2,MIP-1β,TNF-α,IL-1Rα,IL-9,IL-15,FGF-basic,Eotaxin/CCL11,IP-10/CXCL10,MIP-1α/CCL3,PDGF-BB,RANTES,VEGF-A |
| 小鼠趨化因子-31因子Panel | LXLBM31-1 | Luminex | BCA-1/CXCL13,CTACK/CCL27,ENA-78/CXCL5,Eotaxin/CCL11,Eotaxin-2/CCL24,Fractalkine/CX3CL1,GM-CSF,I-309/CCL1,IFN-γ,IL-1β,IL-2,IL-4,IL-6,IL-10,IL-16,IP-10/CXCL10,I-TAC/CXCL11,KC/CXCL1,MCP-1/CCL2,MCP-3/CCL7,MCP-5/CCL12,MDC/CCL22,MIP-1α/CCL3,MIP-1β/CCL4,MIP-3α/CCL20,MIP-3β/CCL19,RANTES/CCL5,SCYB16/CXCL16,SDF-1α/CXCL12,TARC/CCL17,TNF-α |
| 人炎癥37因子Panel | LXLBH37-1 | Luminex | APRIL/TNFSF13,BAFF/TNFSF13B,sCD30/TNFRSF8,sCD163,Chitinase 3-like 1,gp130/sIL-6Rβ,IFN-α2,IFN-β,IFN-γ,IL-2,sIL-6Rα,IL-8/CXCL8,IL-10,IL-11,IL-12(p40),IL-12(p70),IL-19,IL-20,IL-22,IL-26,IL-27(p28),IL-28A/IFN-λ2,IL-29/IFN-λ1,IL-32,IL-34,IL-35,LIGHT/TNFSF14,MMP-1,MMP-2,MMP-3,Osteocalcin,Osteopontin,Pentraxin-3,TNF-R1,TNF-R2,TSLP,TWEAK/TNFSF12 |
| 人趨化因子/細(xì)胞因子-40因子Panel | LXLBH40-1 | Luminex | 6Ckine/CCL21,BCA-1/CXCL13,CTACK/CCL27,ENA-78/CXCL5,Eotaxin/CCL11,Eotaxin-2/CCL24,Eotaxin-3/CCL26,Fractalkine/CX3CL1,GCP-2/CXCL6,GM-CSF,GRO-α (Gro-a/KC/CXCL1),Gro-β/CXCL2,I-309/CCL1,IFN-γ,IL-1β,IL-2,IL-4,IL-6,IL-8/CXCL8,IL-10,IL-16,IP-10/CXCL10,I-TAC/CXCL11,MCP-1/CCL2,MCP-2/CCL8,MCP-3/CCL7,MCP-4/CCL13,MDC/CCL22,MIF,MIG/CXCL9,MIP-1α/CCL3,MIP-1δ/CCL15,MIP-3α/CCL20,MIP-3β/CCL19,MPIF-1/CCL23,SCYB16/CXCL16,SDF-1α+β/CXCL12,TARC/CCL17,TECK/CCL25,TNF-α |
| 人細(xì)胞因子-46因子Panel | LXLRH46-1 | Luminex | CCL2/JE/MCP-1,CCL3/MIP-1 alpha,CCL4/MIP-1 beta,CCL5/RANTES,CCL11/Eotaxin,CCL19/MIP-3 beta,CCL20/MIP-3 alpha,CD40 Ligand/TNFSF5,CXCL1/GRO alpha/KC/CINC-1,CXCL2/GRO beta/MIP-2/CINC-3,CXCL10/IP-10/CRG-2,EGF,FGF basic/FGF2/bFGF,Flt-3 Ligand/FLT3L,G-CSF,GM-CSF,Granzyme B,IFN-alpha 2/IFNA2,IFN-beta,IFN-gamma,IL-1 alpha/IL-1F1,IL-1 beta/IL-1F2,IL-1ra/IL-1F3,IL-2,IL-3,IL-4,IL-5,IL-6,IL-7,IL-8/CXCL8,IL-9,IL-10,IL-12 p70,IL-13,IL-15,IL-17/IL-17A,IL-17E/IL-25,IL-33,Lymphotoxin-alpha/TNF-beta,PD-L1/B7-H1,PDGF-AA,PDGF-AB/BB,TGF-alpha,TNF-alpha,TRAIL/TNFSF10,VEGF |
| 人細(xì)胞因子-48因子Panel | LXLBH48-1 | Luminex | β-NGF,CTACK/CCL27,Eotaxin/CCL11,FGF-basic,G-CSF,GM-CSF,GRO-α (Gro-a/KC/CXCL1),HGF,IFN-α2,IFN-γ,IL-1α,IL-1Rα,IL-2Rα,IL-1β,IL-2,IL-3,IL-4,IL-5,IL-6,IL-7,IL-8/CXCL8,IL-9,IL-10,IL-12(p40),IL-12(p70),IL-13,IL-15,IL-16,IL-17A,IL-18,IP-10/CXCL10,LIF,M-CSF,MCP-1/CCL2,MCP-3/CCL7,MIG,MIP-1α/CCL3,MIP-1β,MIF,PDGF-BB,RANTES,SCF,SCGF-β,SDF-1α,TRAIL,TNF-α,TNF-β,VEGF-A |
| 人炎癥9因子超靈敏Panel | LXMH09-2 | MSD | IFN-γ,IL-1β,IL-2,IL-4,IL-6,IL-10,IL-12p70,IL-17A,TNF-α |
樂(lè)備實(shí)是國(guó)內(nèi)專注于提供高質(zhì)量蛋白檢測(cè)以及組學(xué)分析服務(wù)的實(shí)驗(yàn)服務(wù)專家,自2018年成立以來(lái),樂(lè)備實(shí)不斷尋求突破,公司的服務(wù)技術(shù)平臺(tái)已擴(kuò)展到單細(xì)胞測(cè)序、空間多組學(xué)、流式檢測(cè)、超敏電化學(xué)發(fā)光、Luminex多因子檢測(cè)、抗體芯片、PCR Array、ELISA、Elispot、PLA蛋白互作、多色免疫組化、DSP空間多組學(xué)等30多個(gè),建立起了一套涵蓋基因、蛋白、細(xì)胞以及組織水平實(shí)驗(yàn)的完整檢測(cè)體系。

- Hepcidin-25的作用機(jī)制及在炎癥、心血管與腫瘤等動(dòng)物模型中的應(yīng)用
- Klotho蛋白的分子形式、功能及其在疾病與衰老進(jìn)程中的核心作用
- 二甲雙胍(Metformin)在代謝、腫瘤、衰老與神經(jīng)疾病動(dòng)物模型中的作用
- Romidepsin作為HDAC1/2抑制劑在細(xì)胞分化與損傷修復(fù)中的作用
- 白細(xì)胞介素-2(IL-2)的功能、信號(hào)調(diào)控機(jī)制及對(duì)免疫穩(wěn)態(tài)調(diào)控的雙面性
- 腫瘤/纖維化EMT研究中mIHC染色指標(biāo)的組合及選擇方法
- Hepcidin-25(鐵調(diào)素)在鐵代謝與免疫反應(yīng)中的調(diào)節(jié)機(jī)制
- Vadimezan作為多功能STING激動(dòng)劑與血管破壞劑的作用機(jī)制



