
3Brain高分辨3D電極在腦類器官、神經小球電生理記錄中的應用
如今,有多種源自人類和動物的3D模型,如組織切片、球體、類器官等,能夠以更高的準確性和相關性重現大腦結構和功能的關鍵特征,其效果優于傳統的二維培養。然而,這些3D系統的復雜性不斷增加,帶來了新的方法學挑戰。從生理學角度來看,一個關鍵問題是能否以足夠的時空分辨率探測這些相互連接的內部結構,從而在對生物系統造成最小干擾的情況下準確描述其復雜的網絡活動。
最有前景的方法之一是微電極陣列(MEA),這是一種細胞電子生物接口技術,廣泛用于在體外和離體條件下對神經樣本進行非侵入、無標記及多位點的電生理記錄。除了用于原代細胞或誘導多能干細胞(iPSC)衍生的2D細胞培養物外,這些設備還用于結構化的3D模型,如急性腦片、腦類器官和神經小球等3D培養模型。然而,在3D模型中使用這種技術時,會出現一些關鍵的局限性(圖1):第一個主要的問題在于,傳統的MEA大多數都是平面的,這導致所記錄的信號主要來自最外層。腦片由于制備中的震蕩切片過程,導致其表面神經存活性較差;而對于腦類器官,研究者無法通過平面電極探尋其內部的電活動或者是復雜的神經震蕩。
第二個局限性是,由于3D樣本具備立體結構的特殊性,傳統的MEA不具備足夠的采樣電極數量,去精確捕獲其復雜而精細的電活動。3Brain基于CMOS技術設計的高分辨MEA(HD-MEAs),能夠在一塊芯片上集成數千個電極,從而具備了細胞或亞細胞的空間分辨率,以高時空分辨率的視角,使復雜的神經網絡電動的一覽無余。
第三個問題是應對3D模型中細胞緊密的排列,這需要高效的營養物質擴散、適當的氧氣供應以及快速清除代謝廢物,以避免組織核心迅速壞死。傳統的或平面高分辨微電極陣列(HD-MEA)設備需要細胞與電極緊密接觸才能獲取信號,這意味著組織必須很好地貼附在平面電極上,這一局限性極大的影響了樣本的活性以及快速給藥實驗。3Brain的3D電極設計集成了微流控通道從根本上改善了這些問題,實現長時間維持樣本活性,提高了數據的可靠性。

圖1 2D及3D HD-MEA的對比
在本文中,研究者利用新穎的3D高分辨微電極陣列(3D HD-MEA)技術,對包括急性腦片、腦類器官和神經小球在內的3D生物樣本進行了電生理記錄。首先,研究者展示了電極穿透組織的能力以及微流控通道在提高組織存活率方面的有效性。然后,研究者評估了3D HD-MEA在檢測自發和化學調節活動以及誘發神經活動方面的有效性。總而言之,3D HD-MEA在3D模型中對神經環路特性的生理學研究展現出了巨大潛力。接下來,小編慢慢帶您了解3D電極的方方面面。
在芯片上,研究者實現了兩種規格的3D電極:一種較粗,寬 26um,高 90 um(圖 2A);另一種較細,寬 14um,高 65 um(圖 2B)。電極根部由聚對二甲苯包裹絕緣,僅在針尖處留有可進行電化學反應的界面(圖 2C)。
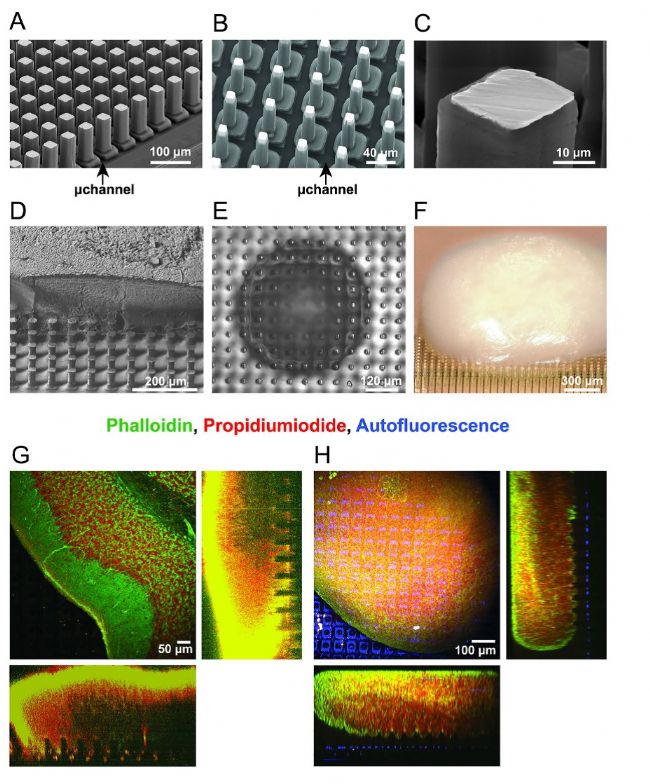
圖 2. 3D分辨微電極陣列芯片的特性及組織穿透情況。A、B)具有較大(a)或較小(b)電極的 3D高分辨微電極陣列的SEM顯微圖像。箭頭指示微流控通道。C)單個3D電極的放大視圖。D)固定在 3D高分辨微電極陣列上的 250 um厚的小腦切片的SEM顯微圖像。用聚焦離子束進行縱向切割,顯示電極穿透組織而未發生彎曲或變形。E)相位對比圖像顯示了放置在3D 高分辨微電極陣列上的腦類器官球體,其中組織與電極的緊密接觸狀態清晰可見。請注意,圖像聚焦在電極尖端所在的平面。F)數字顯微圖像(100x),展示了置于3D 高分辨微電極陣列上的皮層腦類器官。G、H)腦切片(G)和腦球體(H)的雙光子顯微鏡體積掃描圖像,用鬼筆環肽(綠色)和碘化丙啶(紅色)染色,芯片的自發熒光呈藍色。可以觀察到3D電極對組織的穿透情況。
為了測試3D高分辨微電極陣列(HD-MEA)的微流控通道是否能提高組織的存活率,研究者對三種不同的配置進行了活力和功能測試。在連續灌注含氧的克氏液一小時后,將切片從芯片或蓋玻片上輕輕取出,并用 20 µM 的鈣黃綠素AM進行染色,這是一種在活細胞中被酯酶水解后會發出熒光的化合物。從每片切片的兩側獲取共聚焦顯微鏡圖像,從而對鈣黃綠素染色進行定量(圖 3A)。
將切片下側(與芯片或蓋玻片接觸的一側)的活細胞數量與上側(自由表面)的活細胞數量進行了比較。與上側相比,放置在蓋玻片上的切片下側的活力為 38 ± 4%,而放置在 LMC 3D HD- MEA 芯片上的切片下側的活力為 76 ± 3%(圖 3A)。這種差異與下側的接觸面積相符,3D 芯片上的接觸面積為3.6 mm2,而平面芯片上的接觸面積為 15mm2(圖 3B)。這些結果表明,微流控通道能夠提高組織活力,可能是因為改善了切片下層的營養物質擴散和氧氣的交換。
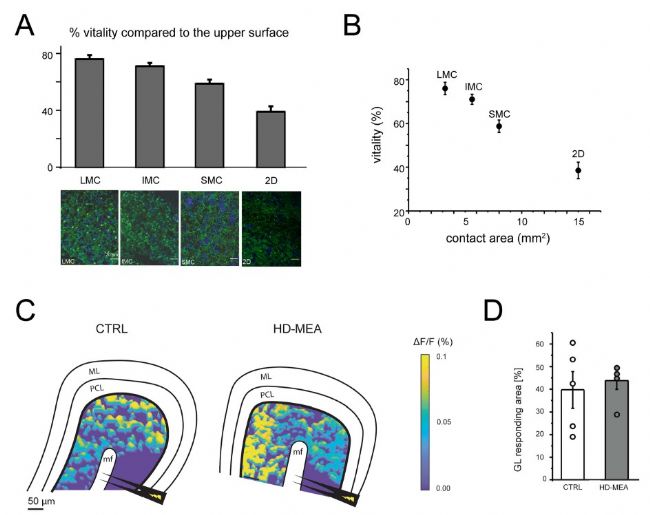
圖3. 平面和3D高分辨微電極陣列(HD-MEA)上小腦切片的組織活力和神經活性。A)直方圖顯示了在平面(2D)或 3D HD-MEA 芯片上放置 1 小時后,小腦切片下表面(與芯片接觸)與上表面(與培養基接觸)的活力對比。下方展示了在 40 倍放大下,用鈣黃綠素 AM 負載的小腦切片在 LMC、IMC、SMC 和 2D 上的代表性共聚焦圖像。活細胞呈綠色;細胞核呈藍色(DAPI)。比例尺 = 20 um。注意 3D HD-MEA 芯片上活細胞的百分比更高。B)該圖展示了 A 中 每種配置下下表面與上表面活力的百分比對比。SMC、IMC 和 LMC 的接觸面積根據微通道寬度估算得出。C)用 VSDi 記錄獲得的偽彩色圖示例,展示了對照組(左)和 3D HD-MEA 上的切片(右)對苔蘚纖維刺激的顆粒層反應的空間分布。刺激電極(黑色)位于苔蘚纖維束(MF)上方。GL,顆粒層。PCL,浦 肯野細胞層。ML,分子層。D)直方圖顯示了在 VSDi 實驗中對苔蘚纖維刺激有反應的顆粒層面積的百分比。所測試的兩種條件下的面積無顯著差異(數據以平均值±均方誤差的形式報告。如文中所述,LMC、IMC、SMC 分別代表具有大、中、小微流控通道的3D高分辨微電極陣列芯片,而 CTRL 表示將組織置于載玻片上。
在急性腦片中檢測3D電極對Spike活動的記錄能力
采用小腦和皮層海馬切片來表征高分辨微電極陣列(HD-MEA)芯片上神經的自發活動。首先將切片放置在平面2D HD-MEA芯片上,然后轉移到3D HD-MEA 芯片上(反之亦然),每次記錄 3 分鐘。與平面HD-MEA芯片相比,3D HD-MEA芯片在檢測小腦浦肯野細胞層的活動方面表現更優,如圖4A中的放電率圖所示。使用 3D HD-MEA記錄的浦肯野細胞層的電活動熱圖與小腦切片的解剖結構匹配度更高,且細節更豐富(圖 4A)。
在小腦切片中,與平面 HD-MEA 芯片相比,在 3D 芯片上進行的記錄顯示出更多的活躍電極(80 ± 19%,2D 上為 390 ± 49 個,3D 上為 790 ± 62 個;n = 6 個切片,p = 0.0006)、更多的神經元個體放電信號(102 ± 23%,2D 上為 371 ± 45 個,3D 上為 668 ± 39 個;n = 6 個切片,p = 0.0005)以及更高的細胞與電極比例(12 ± 5%,2D 上為 1.05 ± 0.01,3D 上為1.18 ± 0.03;n = 6 個切片,p = 0.0126)(圖 4B)。此外,在進行Spike檢測和分類后,與使用平面高分辨微電極陣列記錄的結果相比,3D芯片上的小腦切片中的活躍電極顯示出顯著更高的平均放電率(105 ± 30%;2D上為 25 ± 1.8 Hz,3D 上為 51 ± 5.4 Hz;p = 0.0010),同時峰峰值幅度也有顯著提高(46 ± 14%;2D 上為 76 ± 5 uV,3D 上為 111 ± 9 uV;p = 0.0060)(圖 4C)。

圖4. 3D高分辨微電極陣列(HD-MEA)與平面2D HD-MEA 在小腦切片中的記錄能力比較。A)小腦切片圖像,疊加了由 HD-MEA 電極檢測到的平均放電率的相應電活動圖(顏色刻度條:0 - 10 次/秒;平均放電率 > 0.5 次/秒)。左:切片置于平面 HD-MEA 芯片上;右:切片置于 3D HD-MEA 芯片上。插圖展示了平面和 3D HD-MEA 芯片的掃描電子顯微鏡圖像。B)直方圖顯示了平面和 3D HD-MEA 芯片上記錄到至少一個細胞外單位的電極數量、記錄到的細胞數量以及細胞/電極比率。C)直方圖顯示了平面和 3D HD-MEA 芯片記錄到的單位的平均放電率和峰峰值幅度。在 B 和 C 中,直方圖報告了平均值 ± 標準誤,星號表示顯著差異,* p < 0.05,** p < 0.01,*** p < 0.001。
同樣,使用3D分辨微電極陣列(HD-MEA)檢測到的皮層-海馬切片中的神經元放電活動比平面HD-MEA 更為明顯(圖5A)。與平面HD-MEA 相比,3D HD-MEA 在皮層區域顯示出更多的活躍電極(92 ± 85%,2D 上為56 ± 17 個,3D 上為154 ± 25 個;n = 3 個切片,p = 0.0384)(圖5C)。盡管增加數量沒有統計學意義,但放置在3D HD-MEA 上的切片中的細胞數量(圖5B、C)和檢測到的放電次數(圖5C)均高于平面HD-MEA 芯片上的切片。值得注意的是,3D HD-MEA 檢測到的Spike峰峰值幅度顯著大于平面2D電極(105% ± 30%,2D 上為166 ± 7,3D 上為257 ± 30;n = 3 個切片,p = 0.0221)(圖5C)。
鑒于皮層活動具有“隨機”且稀疏的放電模式,電極可能能夠觸及內層,那里的神經元保存得更好,從而增加了可檢測到的活躍電極的數量。總體而言,這些數據表明,3D 高分辨微電極陣列(HD-MEA)的電極顯著提高了所測試腦區電極的記錄能力。
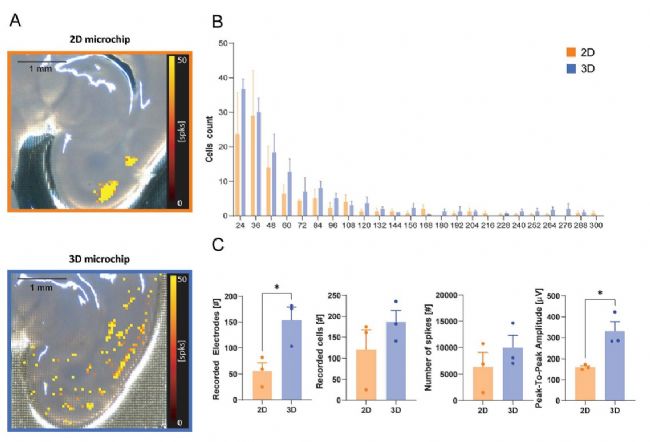
圖 5. 3D高分辨微電極陣列(HD-MEA)相對于平面 HD-MEA 在皮層-海馬切片中的記錄能力。A)皮層-海馬切片的圖像,疊加了由 HD-MEA 電極在 3 分鐘內檢測到的相應電活動圖(顏色刻度條:0 - 50 個Spike)。上:切片放置在平面 HD-MEA 芯片上;下:切片放置在 3D HD-MEA 芯片上。B)平面和 3D HD-MEA 芯片中按細胞計數分布的Spike數量。C)直方圖顯示平面和 3D HD-MEA 芯片記錄的平均活躍電極數量、平均活躍細胞數量、平均總Spike數量以及Spike的峰峰值幅度。在 B 和 C 中,直方圖報告平均值±標準誤,星號表示顯著差異(* p < 0.05)。
加快藥物響應速度
電極底座可防止組織接觸 3D 高分辨微電極陣列芯片的底部,從而在組織下方形成微流控通道網格(圖 2G-H)。除了提高組織的存活率外,底部表面下方溶液的流動性也會得到改善,促進化合物擴散,從而縮短作用時間。為了驗證這一可能性,從放置在平面2D和3D高分辨微電極陣列上的小腦切片中記錄自發活動,分別在灌注含 3 uM河豚毒素(TTX)的克氏液之前和期間進行,時長一分鐘。正如預期的那樣,TTX 灌流逐漸抑制了放電活動(圖 6A)。
然而,放置在 3D高分辨微電極陣列上的切片顯示出更快的抑制速度。在使用TTX處理后的平均放電頻率(神經放電率降低 10%、50% 和 90% 時;所有條件下 p < 0.0001,n = 10 片),其神經元活動完全受到抑制的時間早于平面2D HD-MEA 上的切片(圖 6B 和 6C)。這些結果表明,使用 3D HD-MEA可顯著加快活性化合物的作用,極大的提高實驗效率。
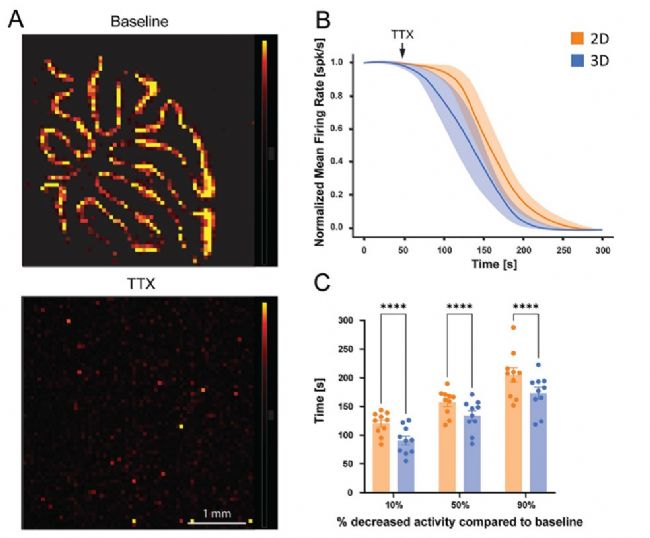
圖6. 3D 高分辨微電極陣列(HD-MEA)與平面 HD-MEA 相比藥物作用加速。為測試藥物灌流的效果,將河豚毒素(TTX)應用于急性小腦切片。A)電活動圖示例,顯示在 3D 芯片上放置的切片在應用 TTX 前(上)和應用后(下)的活躍電極分布情況(顏色刻度條:0 – 300uV)。B)該圖展示了在平面和 3D HD-MEA 上放置的小腦切片在 TTX灌流后活躍單元平均放電率(MFR)的歸一化時間進程。平均放電率歸一化為 TTX 應用前 1 分鐘的基線活動。C)直方圖顯示了兩種條件下 TTX 應用后達到放電頻率降低 10%、50% 或 90% 所需的時間。直方圖報告了平均值 ± 標準誤,星號表示顯著差異,* p < 0.05,** p < 0. 01,*** p < 0.001。
3D電極的電刺激性能
平面2D和3D 高分辨微電極陣列芯片配備了雙向電極,即可用于記錄或者進行電刺激。在小腦切片中,顆粒層對苔蘚纖維刺激的反應速度極快,僅需幾毫秒,因此難以檢測。此外,有效的刺激需要穿透組織以觸及內部苔蘚纖維。基于這些原因,3D電極有望在記錄和刺激誘發的電反應方面發揮關鍵優勢。研究者的結果表明,苔蘚纖維刺激誘發了局部場電位(LFP),該電位在每個受刺激的小葉中傳播(圖 7A),平均傳播速度為 311 ± 39 mm/s(n = 8)。雖然使用 3D 高分辨微電極陣列始終能觀察到快速反應(圖 7A),但同樣的刺激方案在平面高分辨微電極陣列記錄中卻無法激活顆粒層。
此外,研究者還測試了對顆粒層反應的藥理學干預:阻斷 AMPA 受體(10 µM NBQX,圖 7A)和電壓依賴性 Na+ 通道(3 µM TTX,圖 7A)會消除該反應。這種刺激有效地使信號在電路中傳播,這從受刺激小葉浦肯野細胞檢測到的反應中可見一斑(圖 7 B、C)。浦肯野細胞接受顆粒細胞軸突的突觸聯系,表明這種刺激能夠有效地激活神經網絡活動。浦肯野細胞的反應也會因 NBQX 的灌流而消失(圖 7 B、C)。
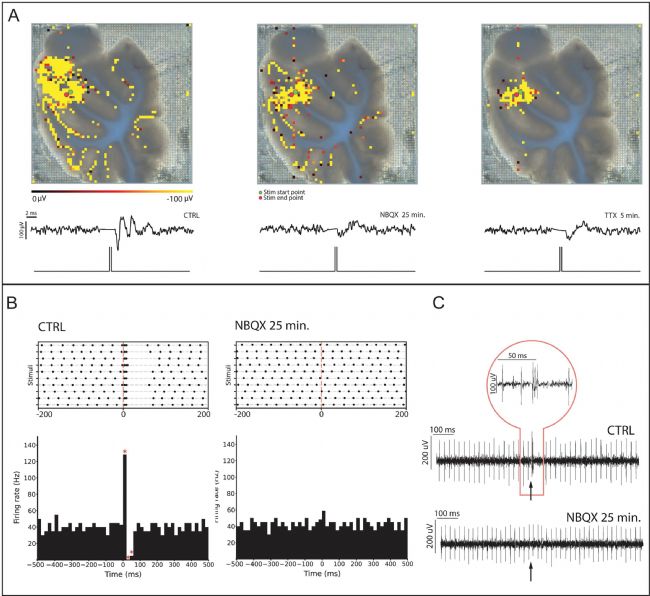
圖 7. 采用3D 高分辨微電極陣列芯片記錄對小腦切片進行電刺激所誘發的神經元活動。A)在苔蘚纖維束對應位置使用雙向電極進行刺激,在不同小葉的顆粒層誘發了反應。由 NBQX 和 TTX 處理后誘發的局部場電位(LFP)消失。電刺激通過小腦切片圖像中紅色和綠色點所指示的通道傳遞,活動圖疊加在圖像上。顏色刻度條:0 - 300 uV。單條曲線分別代表對照條件、NBQX 灌注 25 分鐘后以及 TTX 灌注 5 分鐘后的 LFP。B)對照條件下(CTRL)和 NBQX 灌注后位于刺激小葉的響應性浦肯野細胞的點陣圖和刺激后時間直方圖(PSTH)。相應的原始信號波形圖見 C)。浦肯野細胞的爆發-暫停反應跟隨顆粒細胞的突觸激活,表明刺激有效傳播至網絡。
在具有挑戰性的樣本上測試 3D 高分辨微電極陣列(HD-MEA)記錄效率-PFC自發網絡活動記錄
前額葉皮層(PFC)的神經環路活動與高級腦功能相關,有眾多的科學家渴望記錄該腦區的電活動。但由于記錄 PFC 自發活動的條件本身就很困難,這一局限性極大的限制了對這個腦區的研究。為了測試 3D 芯片增強的記錄能力是否有助于解決這一問題,研究者首先在生理條件下(即不使用改良的細胞外溶液來提高神經元興奮性)記錄了急性切片中前額葉皮層的腹內側區(PrL)的自發活動。研究者在 7 個切片中記錄了 983 個通道顯示基礎自發活動(> 0.1 Hz),占對 PrL 進行采樣的 2284 個電極的 43%。這些單元的平均放電頻率為 0.71 ± 0.09 Hz。為了進一步表征前扣帶回(PrL)的活動,研究者通過灌流一種改良的腦脊液(mACSF),其中不含Mg2+,并增加了K+的濃度,來改變神經元的興奮性,隨后加入抑制性突觸傳遞阻滯劑(10 µM 戊巴比妥)(圖 8A)。
正如預期的那樣,這兩種調整都增加了網絡的興奮性。有趣的是,研究者還觀察到沿著皮質傳播的自發大規模事件信號,表現為局部場電位(LFP)疊加著Spike爆發(圖8B)。這種現象本質上是神經網絡同步活動事件。因此,研究者對兩組切片(有或無傳播的 LFP)進行了表征,以檢測可能存在的基礎活動差異,這些差異或許可以解釋為何在灌注戊巴比妥后,只有部分切片出現了陣發性事件。在缺乏 LFP 的切片組(n = 3;以下稱為組 1)和顯示 LFP 的切片組(n = 4;組 2)之間,基礎放電特性沒有可檢測到的差異。在生理條件下,放電頻率(組 1:363 個單位,0.63 ± 0.07 Hz;組 2:620 個單位,0.79 ± 0.12 Hz; t 檢驗 p = 0.25)

圖 8. 利用 3D 高分辨微電極陣列對小鼠前扣帶回皮質(PrL)電活動的表征。A)示例腦切片,展示了置于 3D 高分辨微電極陣列芯片上的局部場電位(LFP)。從左至右:整個切片,指示 PrL 位置;PrL 區域的放大圖,顯示了在克氏液、改良人工腦脊液(mACSF)以及加巴噴丁灌注期間的連接圖。每種條件下的 3 分鐘自發活動柵格圖顯示在下方。B)來自 A 中同一切片在每種條件(克氏液、改良人工腦脊液和加巴噴丁)下自發活動記錄的原始跡線示例。點為尖峰標記。帶有尖峰的自發 LFP 用條形表示。C)圖中顯示了兩組切片在克氏液中的相關指數(CI)(左),以及第 1 組(中)和第 2 組(右)在測試的三種條件下的 CI 百分比變化。數據以平均值 ± 標準誤(MSE)的形式報告。星號表示顯著差異:* p < 0.05,** p < 0.01,*** p < 0.001。
利用 3D 高分辨微電極陣列測量腦類球體的活動
生成能夠維持三維結構的體外細胞培養物的需求是一項挑戰,近年來,這一需求促使人們開發出了越來越復雜的模型,如類球體和類器官。然而,這類模型通常比體外組織小,細胞密度也更低。鑒于這些原因,研究者采用了更細的 3D 高分辨微電極陣列(HD-MEA)以減少針刺可能造成的損傷(圖 2B)。這些芯片在直徑約為 400 微米和 600 微米的兩種大小的原代胚胎神經元腦類球體上進行了測試。記錄區域(3.8×3.8 mm2))允許在同一個芯片上放置多個樣本,同時由于電極密度高,仍能保持良好的空間分辨率。將不同大小(大、小)的三個類球體安裝在兩個不同的芯片上(圖 9A 左側面板),放置后立即顯示出活動。急性測量顯示存在強烈的尖峰和爆發活動(圖 9A 右側面板)。盡管三個相鄰的球體共用相同的培養基,但它們的活動并未同步,這在圖9A(中間面板)的柵格圖中顯而易見。因此,研究者將每個樣本視為彼此獨立的。較大的球體被更多的電極穿透,因此顯示出更多的電極檢測到活躍細胞(圖 9B)。
然而,無論是大球體還是小球體,“活躍細胞/覆蓋電極”的比例都相似(約 1.5),這表明樣本大小并未直接影響電極的傳感/穿透能力。從每個球體記錄中提取的數據被平均以生成按球體大小排序的實驗亞組。兩組的平均放電率、爆發率和峰峰值幅度如圖 9C 所示。有趣的是,較大的球體顯示出平均放電率和爆發活動增加的趨勢,盡管這在統計學上并不顯著,而峰峰值幅度則不然(圖 9C)。這表明球體大小并未影響信號幅度或神經元-電極耦合。
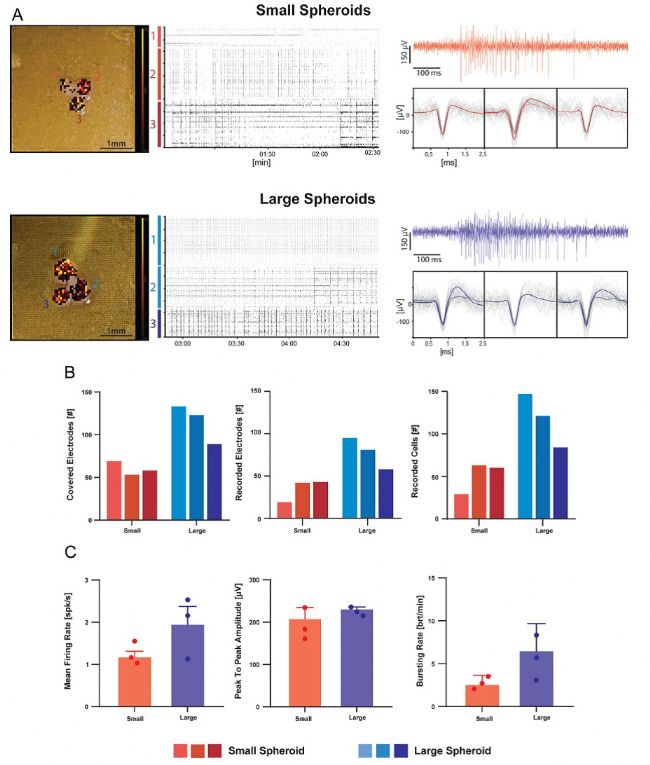
圖 9. 利用 3D 高分辨微電極陣列記錄小鼠腦類器官的電活動。使用了配備更細電極(寬度 14 微米,高度 65 微米)的 3D 高分辨微電極陣列對類器官進行檢測。A)芯片上小類器官和大類器官的圖片與相應的電活動圖(左)疊加,該圖顯示了 3D 高分辨微電極陣列電極檢測到的平均放電頻率(顏色標度條:0 - 10 次/秒)。中間的柵格圖顯示了 2.5 分鐘內的放電活動。三個電極的代表性原始跡線和平均波形如圖所示。)右側。B)直方圖顯示了對小球體和大球體進行采樣的電極數量、活躍電極數量以及活躍單元數量。C)直方圖顯示了小球體和大球體的平均放電率、峰峰值以及爆發率。請注意,球體大小并未影響峰峰值。誤差線表示標準誤差。
利用 3D 高分辨微電極陣列對類器官活性進行化學調節
盡管腦球體具有三維結構,但其細胞組織結構缺乏典型的自組織發育特征,這導致細胞組成和神經回路的復雜性嚴重不足。為克服這些局限性,腦類器官作為一種新型生物學模型應運而生,用于研究人類大腦發育及疾病。將分別由人類誘導多能干細胞(iPSC)培養 219 天和 131 天的皮質和脊髓類器官置于3D高分辨微電極陣列(HD-MEA)芯片上,并置于其培養基中。急性檢測顯示,皮層類器官中有數百個活躍位點表現出自發的放電和爆發活動(圖 10)。檢測到的高度同步性(圖 11B 的柵格圖中明顯可見)表明存在網絡激活。通過向培養基中添加 1uM氯化鉀,放電頻率和爆發頻率分別提高了18%和 14%;
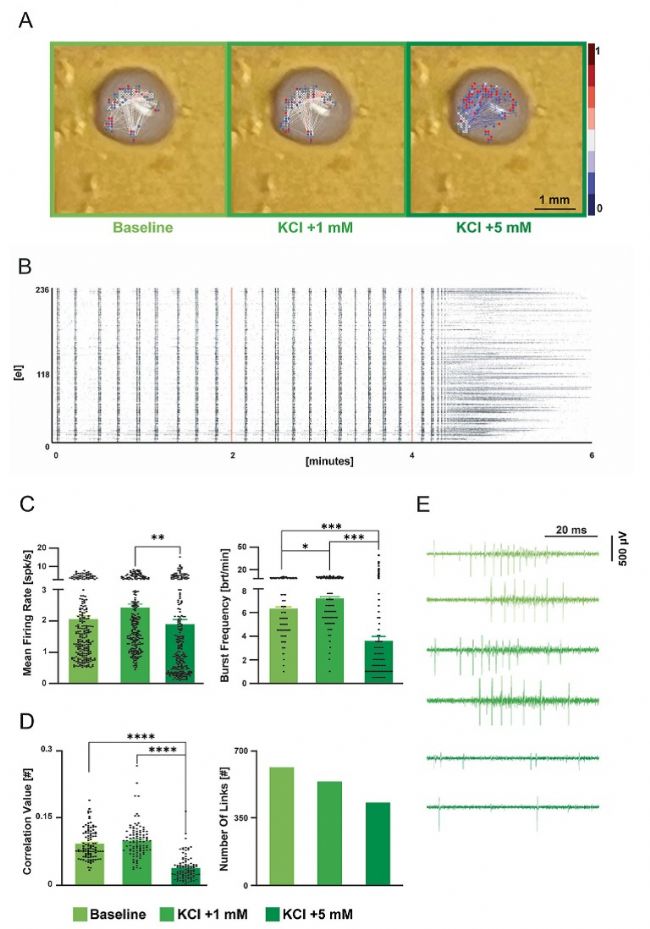
圖 10. 利用 3D 高分辨微電極陣列從人類皮層類器官記錄電活動。A)芯片上類器官的圖片與相應的連接圖重疊(從左至右:基線、+ 1 uM KCL、+ 5 uM KCL)。連接圖是通過互相關分析計算得出。每條連線的顏色根據其歸一化相關程度進行編碼。紅色和藍色點分別代表有入向和出向連接的節點,灰色點代表同時有入向和出向連接的節點。B)表示每種實驗條件下隨時間變化的活動模式的柵格圖。黑色點A)檢測到的尖峰用藍色標記,紅線表示介質中 [K+] 的變化。C)直方圖分別顯示了用于確定活動和網絡同步性的放電和爆發指標。D)直方圖顯示了由互相關分析得出的連接強度和數量。誤差線表示標準誤差。E)每個記錄階段的代表性軌跡,展示了尖峰活動的調制情況。星號表示顯著差異:* p < 0.05,** p < 0.01,*** p < 0.001。
球體和類器官的記錄
3D 高分辨微電極陣列(HD-MEAs)也被證明適用于記錄神經球體和類器官。值得注意的是,腦球體的細胞密度低于腦切片,且體積更小(通常只有幾百微米),這使得在同一樣本中從多個位點進行記錄頗具挑戰性。因此,用于這些記錄的更小的電極尺寸可能至關重要。有趣的是,可以在同一3D HD-MEA 上記錄多個球體,從而有可能同時測試多個樣本。
對于類器官而言,能夠進行急性測量而無需在芯片上培養數天,避免了在平面微電極陣列上常見的細胞遷移效應。這帶來了兩個主要優勢:首先,它形成了由于細胞遷移而形成的二維網絡;其次,它能防止三維結構的改變或壞死核心的增大,這種情況通常會在類器官在自由漂浮環境中生長后被固定時發生。此外,研究者能夠調節皮質和脊髓類器官的放電頻率和同步性,而在這些類器官中進行此類操作通常是困難的。實際上,氯化鉀誘導的膜去極化已知能有效調節脊椎動物和無脊椎動物原代培養物中的神經連接性。
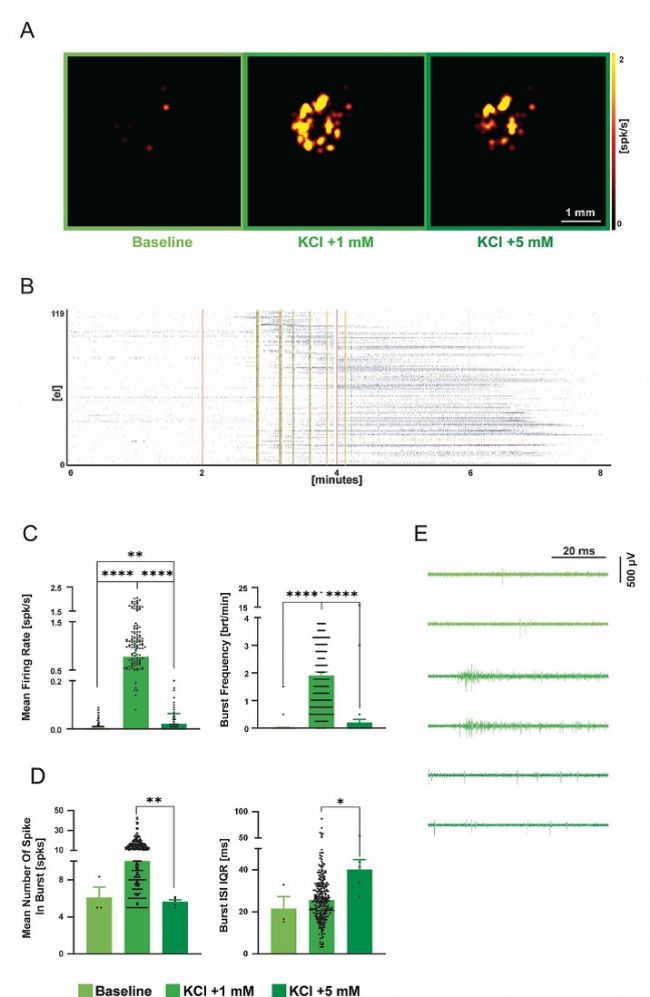
圖 11. 利用 3D 高分辨電極陣列記錄人類脊髓類器官的電活動。A)尖峰圖,顯示了在三種測試條件下芯片上每個電極的平均放電率(從左至右:基線、+ 1 mM、+ 5 mM KCl)。B)柵格圖,顯示了每個實驗條件下的活動模式隨時間的變化。黑色點表示檢測到的Spike。紅色線表示培養基中鉀離子濃度的變化;黃色突出顯示了爆發。C)直方圖顯示了放電和爆發的指標,用于確定活動和網絡同步性。D)提高 KCl 濃度導致1 毫摩爾的氯化鉀(KCl)導致爆發中Spike數量增加,但這種增加并不顯著。相反,更高的氯化鉀濃度(+5 毫摩爾)使爆發中的Spike數量恢復到基線水平(p < 0.0001,n = 230),并增加了爆發Spike間隔的不規則性(p < 0.05,n = 230)。誤差線表示標準誤差。 E)每個記錄階段的代表性軌跡顯示了脈沖活動的調節情況。星號表示顯著差異,* p < 0.05,** p < 0.01,**** p < 0.0001。
結論
3D 高分辨微電極陣列(HD-MEA)技術提高了3D生物模型中電生理記錄和刺激的效率。本文作者認為這項工作為解答此前懸而未決的特定實驗問題奠定了基礎。例如,要繪制腦切片(如大腦皮層、小腦和海馬體)中電信號的分布情況,并解決生理學和計算問題,就需要高分辨率的記錄和刺激位點。3D高分辨微極陣列還為在生理條件下培養和維持的球體和類器官中探索神經活動提供了穩定且可靠的調節,有助于縮小動物組織和人類組織觀察結果之間的差距。在所有這些準備工作中,將使得灌流的藥物產生更快、更徹底的液體和分子交換,提高給藥效率。因此,結合3D高分辨微電極陣列所提供的優勢,研究者將能夠研究神經元活動和突觸可塑性的時空動態以及藥物的神經調節作用。最后,從同一樣本中數千個相互連接的神經元的同步記錄中獲得的大量數據,非常適合用于指導神經網絡的計算模型,從而深入了解神經回路的組織和神經動力學機制。




