
從干細胞到神經細胞——調控子如何 “繪制” 神經系統的多樣性
人類興奮性和抑制性神經元可以通過在體外強制表達先驅轉錄因子(TF)從多能干細胞中誘導產生,這些誘導神經元(iNs)被廣泛應用于研究神經發育、分化以及神經系統疾病(如阿爾茨海默病、帕金森病和肌萎縮側索硬化癥)。盡管轉錄因子可以誘導神經元分化,但生成的神經元亞型多樣性有限,且與體內神經元的功能和轉錄組特征存在差異。
2025年7月10日,蘇黎世聯邦理工學院生物系統科學與工程系Barbara Treutlein教授團隊在Science期刊上發表了題名為“Human neuron subtype programming via single-cell transcriptome-coupled patterning screens”的研究論文,研究試圖探索如何通過組合形態發生素和轉錄因子的協同作用,模擬發育過程中的信號梯度,從而精確控制神經元亞型的命運,豐富體外神經元分化的多樣性。

研究亮點
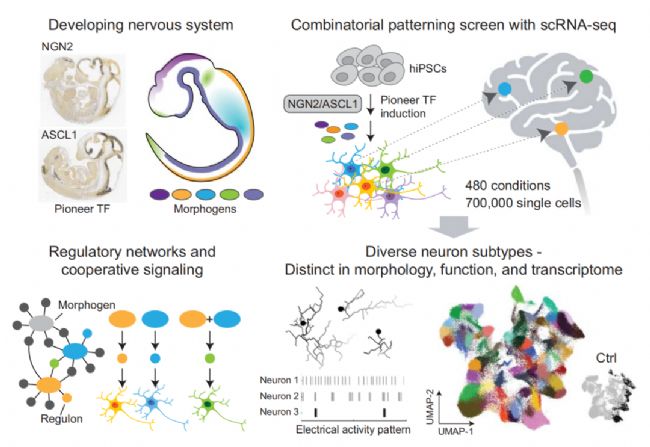
本研究的亮點包括:
1、系統性擴展神經元亞型多樣性
通過 480 種形態發生素組合篩選,結合單細胞轉錄組分析鑒定出涵蓋中樞(前腦、中腦等)和外周神經系統的多種誘導神經元(iN) 亞型。再通過高密度微電極陣列(HD-MEA)分析以及神經遞質釋放分析,揭示不同iN亞型的電活動模式和神經遞質表型具有功能特異性。
2、解析關鍵調控網絡
識別出由轉錄因子及其靶基因組成的 “調控子”(regulons),證實其在形態發生素引導神經元亞型分化中的核心作用,且敲除關鍵轉錄因子會阻斷形態發生素的引導作用,而過表達可獨立驅動亞型形成。
3、優化誘導策略
發現預模式化(先誘導神經前體細胞再用轉錄因子)能產生更均一、更接近原代神經元的 iNs,雖成熟度較低,但轉錄組相似度更高,為精準誘導特定亞型提供新方法。
Maxwell HD-MEA
Maxwell HD-MEA的很多特點使得它受到神經科學家們及人工智能科學家們的青睞:
3265 electrods/mm2的高密度電極
Maxwell MEA芯片上有26400個電極。這樣的密度使其可以記錄2D培養物中幾乎每一個活細胞;而對于3D類器官更為關鍵,因為類器官與芯片接觸面積通常比較小,如此高的密度提供了足夠的記錄位點獲取大量神經元信息。
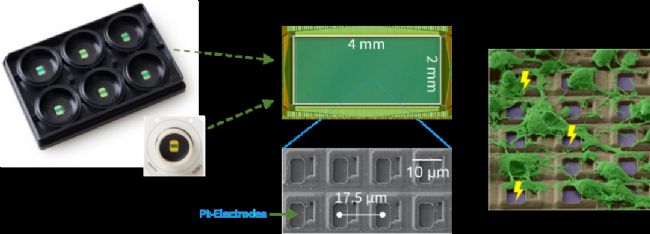
可放在培養箱內進行記錄
這使得在記錄過程中細胞能夠維持良好的生理狀態,支持反復長期的檢測。
低本底噪音,高信噪比
僅為2.4微伏的本底噪音保證了高質量的讀取信號,使得AI系統獲得足夠豐富的輸出信息。
電極可作為刺激電極
在2萬多個電極中每一個電極都可作為刺激電極給出刺激,這在構建的AI系統中成為重要的信息輸入的媒介。在此,高電極密度也為這種信息輸入提供了高空間分辨率的特性。
可開放API,實現快速實時反饋系統
Maxwell HD-MEA可開放API,允許其它軟件的操控,靈活地設計輸入輸出模式,能夠在輸入與輸出間建立實時的反饋。
組合式模式化處理拓展NGN2-iN亞型多樣性
通過組合形態發生素篩選策略,系統探索了NGN2誘導神經元(NGN2-iN)的亞型多樣性。
研究人員設計了192種前后軸(AP)和背腹軸(DV)形態發生素組合,結合單細胞轉錄組測序分析了70萬個細胞,鑒定出101種分子特征明確的神經元亞型。這些亞型不僅覆蓋中樞神經系統各區域(前腦、中腦、后腦和脊髓),還包含多種外周神經元特征。實驗證實形態發生素處理能顯著提升神經元多樣性,如BMP4誘導外周神經標志PRPH表達,而SHH與BMP4在背腹軸模式化中呈現拮抗效應。通過跨物種單細胞圖譜注釋和免疫染色驗證,研究建立了可靠的亞型鑒定體系。批量RNA-seq和時序分析進一步證實該策略具有良好可重復性,產生的神經元亞型轉錄組特征穩定且能持續成熟。
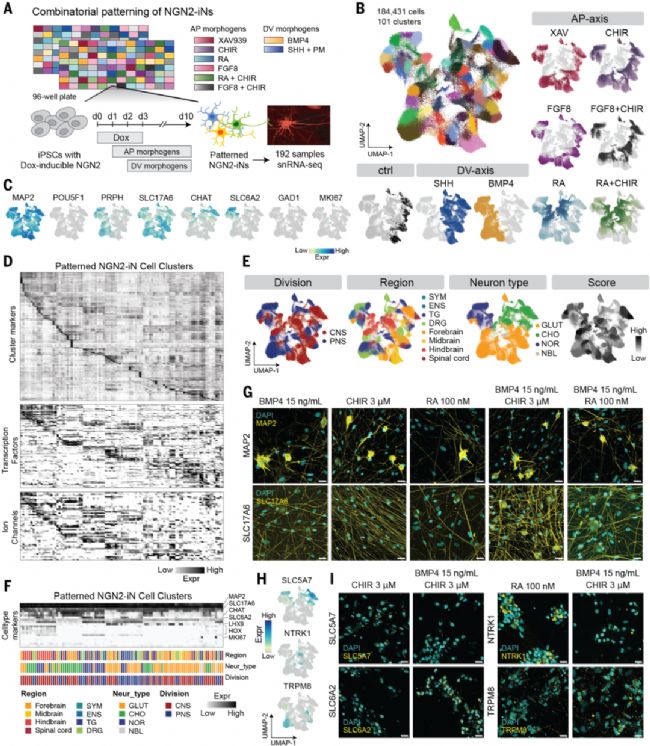
圖1. NGN2-iN神經元亞型組合模式化篩選
(A) NGN2-iN組合模式化篩選實驗設計與時間線,采用單細胞轉錄組測序(snRNA-seq)結合分池組合條形碼技術(Parse Biosciences)分析192種形態發生素組合。(B) 184,431個細胞的UMAP降維圖,按細胞簇(左)和AP/DV形態發生素來源(右)著色。(C) 代表性標志基因表達特征圖。(D) 模式化NGN2-iN細胞簇的標記基因、轉錄因子和離子通道表達熱圖。(E) 基于原代神經元參考圖譜注釋的UMAP圖,顯示神經系統分區(左上)、區域(右上)、神經元類型(左下)和匹配評分(右下)。縮寫:CNS中樞神經系統;PNS外周神經系統;SYM交感神經系統;ENS腸神經系統;TG三叉神經節;DRG背根神經節;GLUT谷氨酸能神經元;CHO膽堿能神經元;NOR去甲腎上腺素能神經元;NBL神經母樣細胞。(F) 模式化NGN2-iN各細胞簇的代表性標志基因熱圖,側邊欄顯示轉移注釋。(G) 第6周模式化NGN2-iN與星形膠質細胞共培養的免疫熒光染色,MAP2(上排)和SLC17A6(下排)呈黃色,DAPI呈青色。比例尺:20μm。(H-I) 第10天模式化NGN2-iN的亞型特異性標志物特征圖(H)和免疫熒光染色(I),包括SLC5A7、SLC6A2、NTRK1和TRPM8。比例尺:20μm。
HD-MEA揭示NGN2-iN的功能與形態多樣性
研究通過高密度微電極陣列(HD-MEA)技術系統分析了模式化 NGN2-iN 的電生理和形態學特征。
選取6種典型條件進行多尺度檢測,發現不同處理組神經元呈現獨特的動作電位模式和放電特性,聚類分析鑒定出11種功能表型。網絡水平分析顯示形態發生素特異性調控神經環路特性,如CHIR處理組具有可重復的電生理特征。藥理學實驗證實AMPA受體拮抗劑可普遍抑制神經活動。軸突重建與Sholl分析揭示不同條件誘導的形態多樣性,且軸突復雜度與轉錄組成熟度高度相關。這些發現證明組合模式化不僅能產生功能成熟的神經元,還可通過精確調控獲得特定電生理和形態特征,為神經回路構建和疾病模型研究提供了重要工具。
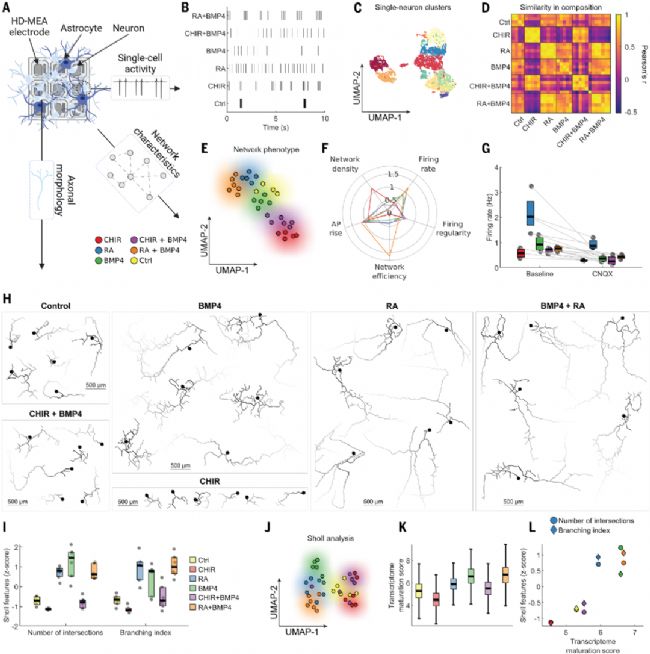
圖2. 模式化NGN2-iN的電生理表征
(A) 星形膠質細胞-NGN2-iN在高密度微電極陣列(HD-MEA)共培養示意圖。HD-MEA可高時空分辨率表征神經網絡活動與軸突形態。電生理記錄于接種后第14天進行(n=66個培養體系)。(B) 六種模式化條件下單個神經元的動作電位序列示例,每條線代表一個鋒電位。(C) 所有推斷的單細胞特征UMAP圖(n=14,997個神經元),按Louvain聚類結果著色。(D) 各培養體系單細胞簇組成相似性熱圖,基于(C)中簇身份計算。(E) 所有單細胞和網絡特征的UMAP圖(n=66個培養體系),按模式化條件著色。(F) 雷達圖顯示五個最重要電生理特征,以對照組為基準歸一化。(G) AMPA/紅藻氨酸受體拮抗劑CNQX處理前后單神經元放電頻率箱線圖(n=43個培養體系)。箱體表示中位數和四分位距,散點為各培養體系平均放電頻率。(H) 基于HD-MEA電信號重建的98個神經元軸突形態,展示每組6個示例。小圓圈示假定胞體/軸突起始段,顏色梯度示動作電位傳導方向。(I) Sholl分析兩個代表性參數的箱線圖(數值經Z標準化),散點代表單個軸突重建結果。(J) 所有Sholl特征的UMAP圖,著色同(I)。(K) 組合篩選scRNA-seq數據中7303個神經元的轉錄組成熟度評分箱線圖。(L) 轉錄組成熟度評分與Sholl特征的散點圖,標記點為各條件均值。
形態發生素組合激活特異性基因調控網絡塑造NGN2-iN神經元多樣性
研究系統解析了形態發生素組合調控NGN2-iN多樣性的分子機制。
通過分析192種處理條件的單細胞數據,發現不同形態發生素具有獨特的調控特性:RA呈現強劑量依賴性,促進后腦膽堿能神經元;BMP4主導外周神經系統特化;而RA+BMP4等組合表現出協同效應。機器學習模型揭示RA和CHIR能引發梯度轉錄響應,而SHH/FGF8的作用較早飽和。
基因調控網絡分析鑒定出形態發生素特異的調控子(regulon):RA激活HOXB4/RARB等后腦決定因子,BMP4通過PHOX2B/MSX1驅動外周神經特征,FGF8相關調控子偏好中腦命運。這些調控子活性與濃度正相關,且在不同細胞系間穩定存在。研究首次在單細胞層面建立了"形態發生素組合-調控網絡激活-神經元命運決定"的完整因果關系,證明組合信號通過協同激活特定轉錄因子網絡實現神經亞型精準調控。
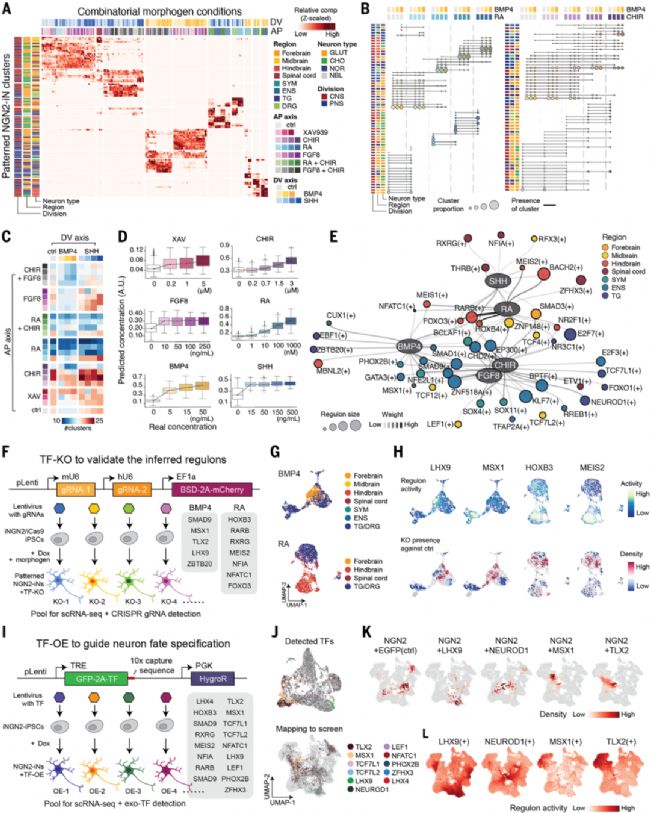
圖3. 組合形態發生素誘導神經元多樣性的調控機制解析
(A) 各組合條件對NGN2-iN細胞簇貢獻的熱圖,細胞數經標準化。(B) 點圖顯示形態發生素組合引起的細胞組成與身份變化,點大小表示簇比例,連線示簇連續性。(C) 各條件生成的細胞簇數量。(D) 隨機森林模型預測的形態發生素濃度與實際濃度對比。(E) 以形態發生素信號調節劑為中心的基因調控網絡(GRN),橢圓節點為形態發生素,圓形節點為調控子(轉錄因子及其靶基因),邊色示關聯強度,節點大小示靶基因數量。(F) 單細胞TF敲除(KO)實驗設計。(G) 基于組合篩選參考注釋的KO細胞UMAP圖。(H) 調控子活性(上)與KO細胞存在度(下)特征圖,藍色示KO后減少。(I) 單細胞TF過表達(OE)實驗設計。(J) TF-OE數據UMAP(上)及其與組合篩選的映射(下),均按外源TF著色。(K-L) TF-OE細胞密度(K)與對應調控子活性(L)特征圖。
模式化NGN2-iN命運決定的形態發生素-GRN調控機制
通過單細胞水平的基因編輯和過表達實驗,驗證了形態發生素通過特定轉錄因子調控網絡(GRNs)決定神經元亞型的機制。
敲除實驗顯示,LHX9、MSX1等關鍵轉錄因子的缺失會特異性消除對應調控子活躍的神經元亞群,如MSX1敲除減少外周神經細胞,HOXB3敲除抑制后腦神經元生成。過表達實驗則證明,NGN2與亞型特異性TF(如PHOX2B等)的組合表達,無需外源形態發生素即可誘導目標神經元亞型,且純度更高。特別發現某些TF(如MSX1)的過表達效果受時序調控影響。這些結果不僅證實了形態發生素-GRN推斷模型的準確性,更建立了"核心TF組合決定神經元命運"的普適性規律,為神經再生醫學提供了精準操控細胞命運的新范式。
預模式化神經祖細胞狀態決定NGN2誘導神經元的獨特身份特征
預模式化指的是在誘導神經分化(如表達NGN2/ASCL1等先驅轉錄因子)之前,先通過形態發生素處理多能干細胞,使其先分化為具有特定區域身份的神經祖細胞(NPCs),再進行神經元誘導。后模式化指的是誘導多能干細胞直接分化為神經元(如通過NGN2過表達),之后再施加形態發生素進行區域特化。
研究通過對比預模式化與后模式化策略,揭示了神經祖細胞初始狀態對終末神經元身份的決定性作用。設計96種預模式化條件結合單細胞多組學分析,發現不同處理會塑造神經祖細胞特異的表觀遺傳:前腦祖細胞富集OTX2結合位點,后腦祖細胞則呈現CDX4調控特征。NGN2作為先鋒因子,通過開啟與神經成熟相關的染色質封閉區域啟動分化。關鍵發現包括:(1)預模式化神經元保留祖細胞的區域身份,證明NGN2主要執行分化而非區域化功能;(2)RA天然誘導后腦祖細胞表達NGN2,揭示內源調控機制;(3)預模式化產生更接近體內發育的神經元亞型,且異質性更低。
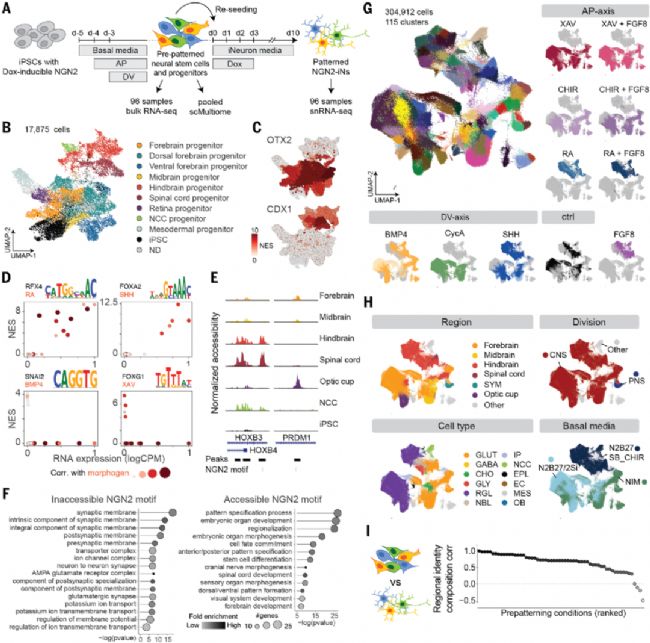
圖4. 多樣化預模式化輸入狀態下的NGN2誘導擴展神經元多樣性
(A) 預模式化篩選與NGN2誘導實驗設計。(B) 預模式化神經干/祖細胞UMAP圖,按注釋著色。(C) 按可及區域基序富集[NES]著色的UMAP圖。(D) 基序富集與TF表達相關性散點圖。(E) NGN2基序可及性差異的ATAC峰示例。(F) NGN2基序可及/不可及關聯基因的GO分析。(G-H) 304,912個預模式化NGN2-iN細胞的UMAP圖,按簇身份/形態發生素來源(G)或原代注釋/基礎培養基(H)著色。(I) 預模式化神經干/祖細胞與NGN2-iN的神經身份組成相關性,每點代表一個形態發生素組合。
預模式化與后模式化NGN2-iN在細胞組成、成熟度及調控子活性上的比較
研究系統比較了預模式化與后模式化NGN2神經元的特征差異。
預模式化產生更接近體內神經元的均質群體,但成熟較慢且含更多未成熟膠質細胞;后模式化神經元成熟更快但異質性更高。關鍵發現包括:(1)預模式化能獲得視網膜前體等特殊亞型;(2)祖細胞的表觀遺傳記憶延緩但優化了分化進程;(3)相同形態發生素在不同時序激活差異化的調控網絡。例如RARB在兩種模式下雖均調控后腦命運,但靶基因譜不同。這些發現不僅揭示了神經分化時序調控的重要性,更證明祖細胞初始狀態如同"發育密碼",持續影響終末神經元的功能特性,為優化體外神經分化體系提供了重要指導。
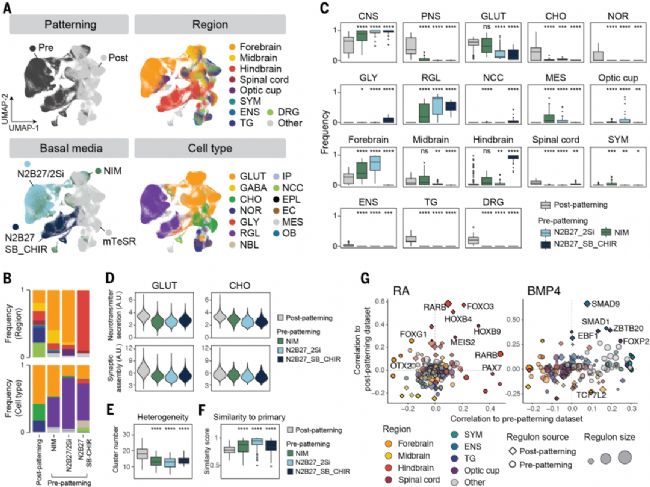
圖5. 預模式化與后模式化NGN2-iN的比較
(A) 整合數據集489,343個細胞的UMAP圖。(B-C) 區域身份(B)和細胞類型(C)頻率組成比較。(D) 基于神經遞質分泌和突觸組裝相關基因的成熟度比較。(E-F) 異質性(E)和原代相似性(F)比較。(G) 預/后模式化調控子與RA/BMP4活性的相關性比較,點大小示調控子規模。
轉錄因子誘導結合組合模式化的普適性細胞編程策略
通過將組合模式化策略拓展至ASCL1-DLX2誘導的抑制性神經元,建立了普適性細胞編程體系。
關鍵發現包括:(1)不同先驅轉錄因子(NGN2/ASCL1-DLX2)結合相同形態發生素組合可產生截然不同的神經元亞型譜系,證明該策略具有廣泛適用性;(2)預模式化雖提高細胞純度但伴隨更多脫靶細胞,揭示DLX2等多效性因子的譜系限制作用;(3)構建的整合神經元圖譜包含185個分子特征明確的亞群,最高純度達84%,為神經科學研究提供豐富資源;(4)發現形態發生素濃度與神經肽表達的定量關系(如CHIR-NPY、BMP4-生長抑素),為疾病模型構建提供調控靶點;(5)系統比較顯示模式化iNs能準確模擬原代神經元的轉錄因子網絡和電生理特征,但在能量代謝方面存在顯著差異,提示體外體系仍需優化。這些發現不僅證實了"轉錄因子+形態發生素"組合策略在神經分化中的普適性,更通過建立表型-條件-調控網絡的定量關系,為再生醫學和疾病建模提供了標準化細胞編程框架。特別值得注意的是,研究揭示了體外培養環境對細胞代謝的重編程效應,這為改進神經元成熟度提供了重要方向。
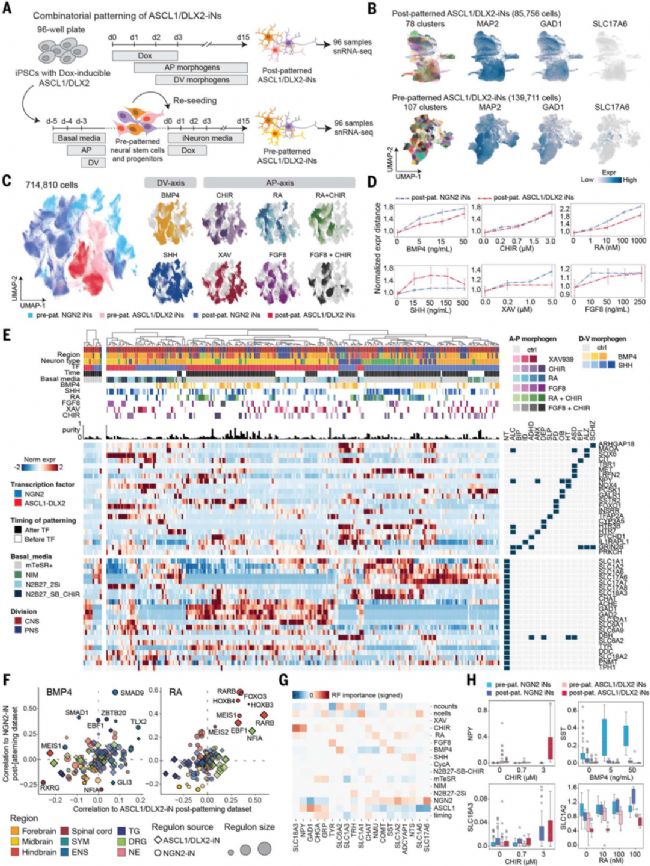
圖6. 組合模式化產生多樣化GABA能ASCL1/DLX2-iN
(A) ASCL1/DLX2誘導的預/后模式化篩選設計。(B) ASCL1/DLX2-iN的UMAP圖。(C) 430,936個神經細胞的整合圖譜UMAP。(D) 形態發生素濃度與轉錄差異的散點圖。(E) 基于轉錄組距離的聚類樹狀圖,熱圖示各簇最佳條件的濃度,條形圖示簇純度。(F) NGN2與ASCL1/DLX2-iN調控子與BMP4/RA相關性比較。(G) 形態發生素對神經遞質/神經肽基因表達的隨機森林預測重要性熱圖。(H) 神經遞質/神經肽表達與形態發生素濃度的箱線圖。
研究總結
這篇研究通過系統篩選480種形態發生素組合,結合單細胞測序、高密度微電極陣列(HD-MEA)等技術,在人類干細胞中建立了定向誘導多樣化神經元亞型的普適性策略。
研究發現預模式化(先區域化后分化)能產生更接近體內神經元的亞型,而后模式化(先分化后區域化)則更快速高效。通過解析形態發生素-轉錄因子調控網絡,研究不僅揭示了神經元命運決定的協同信號機制,還構建了包含400余種亞型的神經元圖譜。該成果為神經系統疾病建模、藥物篩選和細胞治療提供了重要工具和理論框架,是干細胞神經定向分化領域的重大突破。
參考文獻:
Lin HC, Janssens J, Eisinger B, et al. Human neuron subtype programming via single-cell transcriptome-coupled patterning screens. Science. 2025 Jul 10;389(6756):eadn6121. doi: 10.1126/science.adn6121.
想要獲取本篇文獻的老師同學們,可以拉到文末掃碼添加禮智小客服哦!




