
七鰓鰻單細胞轉錄組圖譜揭示Natterin誘導白色脂肪組織的褐變研究

期刊:Nature communications
影響因子:15.7
主要技術:scRNA-seq、snRNA-seq
導語
七鰓鰻作為一種無頜脊椎動物,在理解脊椎動物進化方面扮演著關鍵角色。然而,由于缺乏對多種七鰓鰻器官的細胞學研究,阻礙了該領域的進展。因此,本研究構建了一個包含 604460 個細胞核和 70 種細胞類型的綜合細胞圖譜,涵蓋了 14 個七鰓鰻組織樣本。跨物種的細胞進化比較表明,大多數七鰓鰻細胞類型與有頜脊椎動物的同源。盡管七鰓鰻缺乏實質器官,但我們發現了腺泡樣和胰島樣細胞群,這為脊椎動物胰腺功能的證據提供了依據。此外,我們研究了七鰓鰻免疫細胞群的異質性。Natterin 在顆粒細胞中高度表達,定位于脂滴中。我們還開發了一種表達 Natterin 的轉基因小鼠模型,以闡明 Natterin 在脂質代謝中的作用,從而誘導白色脂肪組織褐變。這些發現闡明了脊椎動物的細胞進化,并推進了我們對七鰓鰻脂肪組織可塑性和代謝調節的理解。
主要技術
scRNA-seq、snRNA-seq
研究結果
1. 通過綜合細胞圖譜揭示的組織與細胞特征
我們從 30 條七鰓鰻幼體中全面采集了 14 種組織,并使用 scRNA-seq 和snRNA-seq 技術進行了單細胞測序和單核測序(圖 1a)。總共獲得了 604460 個細胞/核。從血液中的 130654 個到大腦中的 5813 個,細胞數量在不同組織中有所不同(圖 1b、c)。此外,UMAP揭示了每個組織的不同簇,來自同一系統的組織表現出更緊密的聚類模式(圖 1b、d)。我們使用已報道的標記物和高度表達的基因將這些細胞簇分配到所有組織的全局 UMAP 視圖中的 70 種細胞類型(圖 1d)。因此,我們成功地構建了七鰓鰻的綜合細胞圖譜,這使我們能夠進一步分析所有七鰓鰻組織的細胞特征。
構建的細胞圖譜被用來闡明七鰓鰻組織的細胞特征。腎小球細胞、近端小管、集合管、間質細胞、遠端小管、亨氏環和髓袢細胞等組織特異性細胞僅分布在腎臟中(圖 1d)。同樣,我們在肝臟中觀察到組織特異性細胞,包括膽管細胞和肝細胞,以及肝星狀細胞、庫普弗細胞、造血干細胞和導管細胞(圖 1d)。肝星狀細胞在維生素 A 儲存、細胞外基質產生、纖維化、肝臟再生調節和免疫調節中起著關鍵作用。同樣,庫普弗細胞是專門的巨噬細胞,促進肝臟免疫反應、清除病原體和毒素、維持肝臟組織完整性以及調節肝臟炎癥和免疫穩態。我們的細胞圖譜揭示的細胞組成,提供了對七鰓鰻細胞的詳細且前所未有的圖解。
然后,我們比較了不同組織中的細胞類型。在鑒定的 70 種細胞類型中,包括內皮細胞和各種免疫細胞在內的 22 種細胞類型在不同的組織中被發現(圖 1d)。像心肌細胞和膽管細胞這樣的 48 種細胞類型是組織特異性的。我們使用內皮細胞作為例子來探索這些細胞類型的特征。內皮細胞存在于各種組織中,包括心臟、腎臟、肝臟、尾鰭、上位體、脊索、鰓、背鰭、肌肉、皮膚和大腦(圖 1d)。盡管它們不具有特異性,但內皮細胞在不同組織中有所不同。使用基于GSVA的 439 個血管相關基因的精選集對高度表達的基因進行功能富集分析也反映了組織特異性血管內皮細胞的不同功能。腦內皮細胞中的環核苷酸代謝過程被證明可以調節血管張力、血腦屏障完整性、腦血流量和通透性。相比之下,肝臟內皮細胞中的補體和凝血級聯通路調節補體系統、凝血級聯、止血和炎癥。
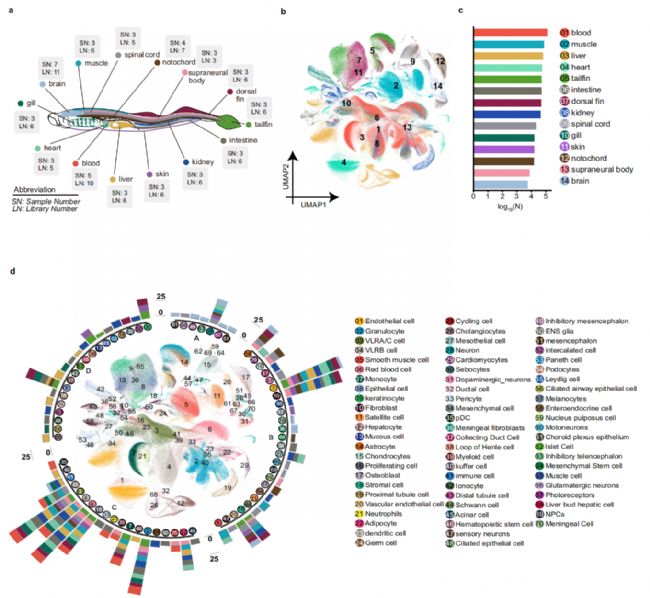
圖1
2. 通過跨物種比較追蹤細胞類型進化方向
細胞類型的鑒定使我們能夠檢查以前未探索的組織和器官。我們根據七鰓鰻物種的可用細胞圖譜將七鰓鰻細胞類型與小鼠和人類細胞類型進行對齊。這三個物種之間的細胞表現出高度相似性,表明存在共同的細胞血統。例如,七鰓鰻上皮細胞表現出與哺乳動物肺泡 II 型細胞非常相似,而七鰓鰻橈側神經膠質細胞表現出與哺乳動物少突膠質細胞的高度相似性。此外,七鰓鰻變異淋巴細胞受體 B (VLRB) 細胞與哺乳動物 B 細胞顯著相似。我們鑒定了 92 個七鰓鰻胰島樣細胞(表達標記基因 Ins、Cpe 和 Epas1)19 和 689 個腺泡樣細胞(表達標記基因 Ctrb1 和 Cel),它們與來自小鼠和人類的胰腺特異性細胞對齊,盡管七鰓鰻缺乏明顯的胰腺。這些細胞定位于腸道(圖 2a),表明七鰓鰻的腸道可能實現的胰腺功能。我們收集了先前報道參與脊椎動物胰腺發育和代謝的基因集,并描述了它們在七鰓鰻胰島樣細胞和腺泡樣細胞中的表達特征,與小鼠和人胰腺細胞相比。標志性胰腺β細胞特異性基因在七鰓鰻胰島樣細胞中特異性表達,而naba基體增生胰島和黑板胰腺白細胞衰老特異性基因在腺泡樣細胞中特異性表達。我們對七鰓鰻腸的橫截面進行了HE染色和FISH,以驗證這些物種中胰腺樣細胞的位置。同樣,通過FISH驗證了腸道中Ctrb1、Cel和Pnliprp2的表達。同樣,免疫組化結果顯示腸道內胰島素的強烈陽性信號,與FISH結果一致(圖 2b)。這些發現進一步表明了七鰓鰻腸道可能的完整胰腺功能,為七鰓鰻器官組織和功能以及脊椎動物器官進化提供了見解。
我們使用細胞圖譜對免疫細胞進行了廣泛的分析,以闡明七鰓鰻的免疫特征和脊椎動物免疫系統的進化。我們圖譜中包含的 280636 個免疫細胞被聚集成不同的細胞類型,包括 68924 個粒細胞、58622 個 VLRA/C 細胞(Vlra 和 Vlrc 在同一簇中表達)、44116 個 VLRB 細胞、30894 個單核細胞、5469 個中性粒細胞、4875 個樹突狀細胞、2197 個 pDC 細胞、1213 個髓系細胞和 1074 個 Kupffer 細胞(圖 2c)。這些細胞類型表現出基于細胞類型和組織起源的聚類模式(圖 2c,d)。然后,我們將七鰓鰻免疫細胞類型與小鼠免疫細胞類型進行比對,可變淋巴細胞受體 (VLR) 細胞是七鰓鰻所獨有的。相比之下,七鰓鰻 VLR 細胞和小鼠 T/B 細胞共享同源基因,包括 Syk、Map3k1 和 Pik3cd。此外,它們的平均表達水平表現出VLR或T/B細胞特有的相似模式,與其他免疫細胞類型相似。這些發現強調了七鰓鰻 VLRB 細胞和脊椎動物 B 細胞之間的相似性,以及七鰓鰻 VLRA/C 細胞與 T 細胞的同源性。此外,VLRB細胞共表達巨噬細胞相關基因,如Csf1r。免疫熒光測定進一步證實了這一觀察結果,表明 VLRB 細胞可能與脊椎動物巨噬細胞具有相似的功能或具有共同的生理特征。這些結果為七鰓鰻的免疫細胞多樣性提供了寶貴的見解,并證明了 VLRB 細胞的潛在功能作用和進化。
粒細胞是七鰓鰻中最豐富的免疫細胞群(圖 2c),包括四種不同的亞型:Thbs1+、Retn+、Haaf+ 和 Aldh1a1+(圖 2e). 主要與炎癥、組織重塑、傷口愈合和病原體反應相關的途徑相關的基因在這些細胞中高度表達。特別是,Retn+粒細胞表現出類似于下頜脊椎動物巨噬細胞的特征,在中性粒細胞脫顆粒、基質金屬蛋白酶和白細胞脫顆粒中表現出功能富集。這些觀察結果有力地支持將這些細胞鑒定為粒細胞。此外,這些發現表明七鰓鰻粒細胞具有血細胞樣細胞和巨噬細胞樣細胞的組合特征,使它們能夠激活免疫反應。
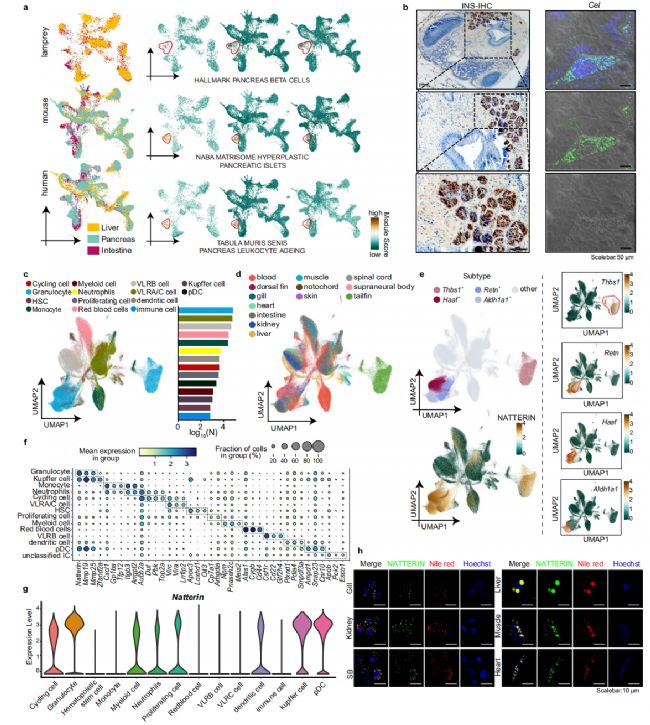
圖2
3. Natterin介導小鼠鐵依賴性 WAT 褐變
Natterin是粒細胞亞群中差異表達最顯著的基因(圖 2f,g)。通過免疫熒光和免疫組織化學進一步表征了Natterin在各種組織中的表達譜,這些結果支持Natterin對脂滴的定位(圖 2h)。Natterin是一種古老的先天免疫相關基因,分布廣泛,并在植物、真菌、無脊椎動物和脊椎動物等各種物種中經歷了獨特的進化。Natterin 存在于多種七鰓鰻、射線鰭魚、爬行動物和鳥類中,但在盲鰻、軟骨魚、肺魚、兩棲動物(Rana temporaria、Pelobates cultripes、Pelobates fuscus 和 Pleurodeles waltl 除外)和哺乳動物(針鼴除外)中不存在,我們預測了六個代表性物種的Natterin結構,在凝集素結構域和哺乳動物特異性螺旋的結構上顯示出顯著差異(圖 3a)。這些發現表明,隨著物種的進化,Natterin 存在不同的進化進程和功能變異。
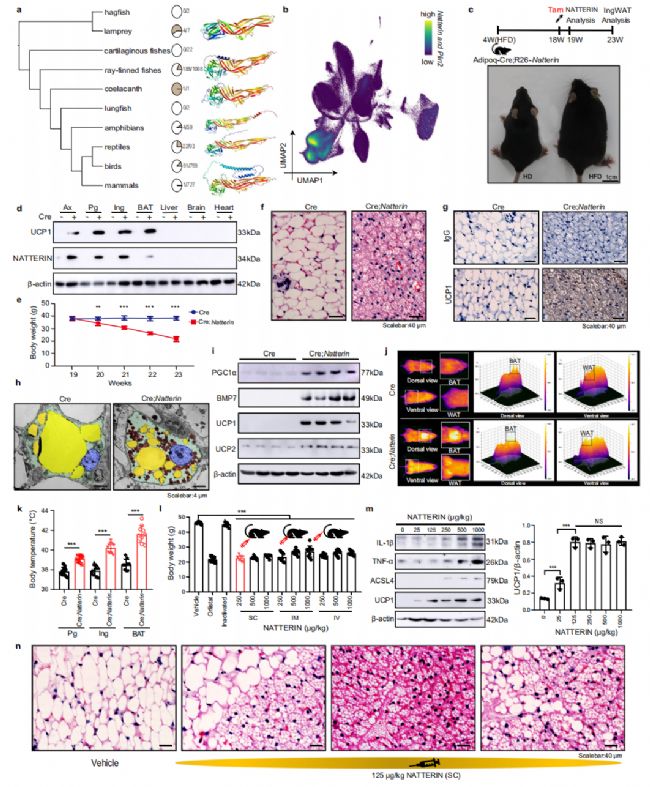
圖3
根據我們的scRNA-seq數據,我們觀察到粒細胞中也表達Plin2基因的Natterin(圖 3b)。雖然 Natterin 被廣泛認為是一種參與宿主防御的免疫分子,但其參與脂質代謝的探索較少。因此,我們從七鰓鰻組織中提取并分析了脂滴(圖 2h)。在七鰓鰻細胞中觀察到類似白色脂肪細胞的大脂滴和類似棕色脂肪細胞的較小多房脂滴。棕色脂肪細胞表現出中央細胞核和高線粒體密度,表明代謝狀態活躍。為了研究 Natterin 在脂質調節中的潛在作用,我們開發了一種轉基因小鼠模型(Cre;Natterin)與飲食誘導的高脂肥胖,以進一步闡明 Natterin 在脂肪組織中的作用。該模型使用 Cre-loxp 系統實現了 Natterin 的脂肪組織特異性表達(圖 3c、d)。在第 18 周,我們給予他莫昔芬以誘導 Cre 中 Natterin 的表達;Natterin 小鼠,隨后在第 19 周評估 Natterin 和 Ucp1 的表達(圖3d)。我們觀察到 Cre 和 Cre 之間的體重存在顯著差異。電鏡顯示Cre;Natterin小鼠表現出分裂成幾個具有高線粒體含量的小多室脂滴(圖 3h)。這些形態變化是WAT褐變的特征。此外,我們檢測到UCP1和幾個與WAT褐變相關的標記基因的高表達,如PGC1α和BMP7(圖 3i),表明 Natterin 可以誘導WAT的褐變。此外,紅外成像和涉及冷暴露和熱中性的實驗表明,在 Natterin 過表達后,脂肪組織(WAT和BAT)的產熱能力顯著增加(圖 3j、k)。這些結果表明, Natterin 在體內誘導脂肪組織產熱。
我們在小鼠身上進行了實驗,使用三種注射方法測試不同劑量的 Natterin 的效果。Natterin 表現出與市售減肥藥奧利司他(對照:46.24 ± 0.68 g,NATTERIN:22.63 ± 1.63 g,奧利司他:21.65 ± 1.65 g)表現出相當的效果,皮下注射是最有效的方法。 Natterin 誘導的體重減輕伴隨著WAT褐變,如WB、HE染色和UCP1、PLIN2和COX IV(線粒體的標記基因)的免疫組織化學分析所證明的那樣(圖 3m,n)。此外,我們發現皮下注射低劑量(125 μg/kg)的 Natterin 不會引起任何器官損傷,引發鐵死亡和炎癥反應(圖 3m)。這表明,使用低劑量 Natterin 的肥胖小鼠體重減輕不是由于它們的免疫功能。
4. Natterin 影響 WAT褐變的潛在細胞機制
為了闡明 Natterin 對WAT影響的潛在細胞機制,我們研究了用這種蛋白質處理后WAT細胞中Fe2+水平的顯著增加(圖 4a)。我們提出 Natterin 介導 WAT 細胞中的細胞內鐵過載。我們使用 Biacore 技術發現了這種蛋白質與轉鐵蛋白受體 (TFR1) 的強大結合能力(圖 4b)。免疫熒光分析顯示,皮下注射 125 μg/kg 重組 Natterin 進行體內和體外研究的小鼠的 WAT 中 Natterin 和 TFR1 的共定位(圖 4c)。結果表明, Natterin 在白色脂肪細胞膜表面大量積累,其中 Natterin(綠色熒光)高度集中在 TFR1 陽性(紅色熒光)區域。ATGL介導的脂肪分解導致脂滴分解,游離脂肪酸的產生可以激活UCP1功能。我們進一步分析了ATGL(脂肪分解標志物)、DRP1(線粒體裂變標志物)和CPT1A(線粒體脂肪酸代謝中的限速酶)響應 Natterin 對TFR1的調節的表達(圖 4d)。這表明 Natterin 在 TFR1 介導鐵依賴性細胞內褐變分子機制中發揮作用。
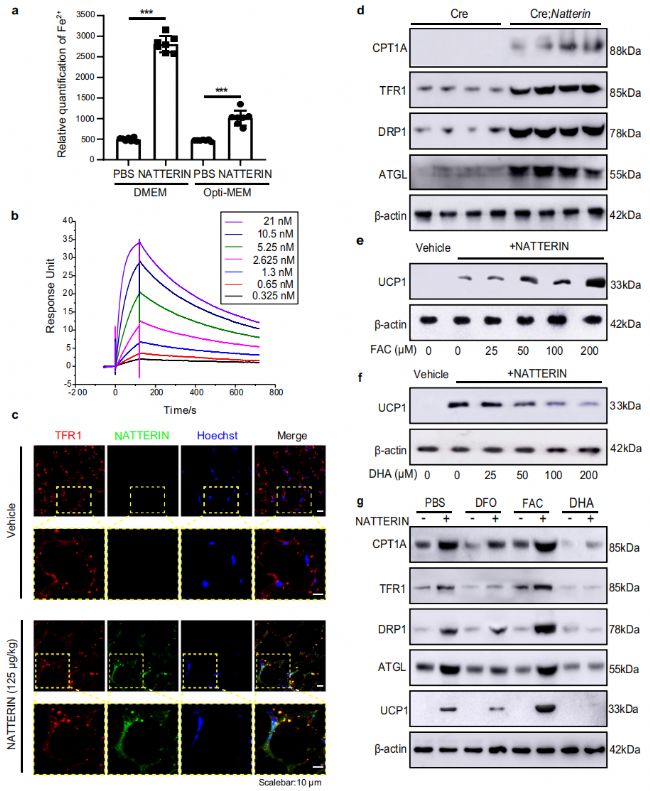
圖4
我們開發了一種用檸檬酸鐵銨(FAC)、去鐵胺(DFO)和二氫青蒿素(DHA)處理的小鼠模型,以評估鐵代謝對WAT細胞的影響(圖 4e,f)。FAC 和 DFO 是分別促進和抑制細胞內鐵含量的鐵調節劑,而 DHA 抑制 TFR1。FAC促進 Natterin 介導的白色脂肪褐變,DHA抑制 Natterin 介導的褐變,同時抑制TFR1,這兩個過程都是劑量依賴性的。我們的結果顯示,在TFR1的正向和負向調控下,這些標記物的表達發生了一致的變化(圖 4g),表明細胞鐵水平的增加促進了ATGL介導的脂肪分解,導致通過白色脂肪細胞的分解形成許多小脂滴。ATGL介導的脂肪分解產生游離脂肪酸,并能激活脂肪細胞線粒體的解偶聯功能。此外,線粒體裂變因子DRP1表達的上調促進了線粒體增殖,最終促進了白色脂肪細胞的褐變。這種褐變過程表明, Natterin 促進了線粒體內膜中 UCP1 的表達,并且 UCP1 介導了未偶聯的氧化磷酸化,以線粒體產生的 ATP 的化學能作為熱量釋放出來。 Natterin 介導線粒體裂變和解偶聯,促進脂肪細胞代謝。這些發現表明, Natterin 通過與 TFR1 相互作用并誘導顯著的代謝重編程來介導 WAT 細胞中的鐵積累。這種重編程包括增強脂肪分解和線粒體生物發生,有助于白色脂肪細胞的褐變并可能改善代謝特征。
結語
在這項研究中,我們使用scRNA-seq和snRNA-seq技術構建了七鰓鰻器官的綜合細胞圖譜。我們進一步比較了七鰓鰻和有頜脊椎動物之間的細胞組成和相似性。七鰓鰻和有頜脊椎動物之間共享幾種同源細胞類型,盡管解剖學存在差異,但功能相似。特別是,我們在七鰓鰻的腸道中鑒定了腺泡樣和胰島樣細胞類型,表明即使在這些生物體沒有胰腺的情況下,也會出現葡萄糖調節功能。胰腺細胞聚集成簇,分散在腸壁和管子與食道的交界處。我們的研究結果闡明了七鰓鰻的進化適應,并為與有頜脊椎動物相比,其細胞組成的功能異同提供了寶貴的見解。盡管已經確定了七鰓鰻和有頜脊椎動物之間的同源細胞類型,但驅動這些相似性的分子機制及其進化意義仍有待充分探索。
除了建立七鰓鰻的細胞景觀外,我們還評估了免疫細胞。我們分類了4種粒細胞亞型,它們不僅富集于免疫相關途徑,而且還具有脂肪細胞分化調節和冷誘導產熱的作用,提示七鰓鰻粒細胞可能參與脂質代謝。 Natterin 在 WAT 褐變中的作用的發現進一步證明了七鰓鰻免疫蛋白的多功能性,并揭示了這些生物體調節其代謝過程的潛在機制。IL-27 通過直接靶向脂肪細胞、促進產熱、增強代謝功能以及提供預防肥胖的保護,在 WAT 褐變中起著至關重要的作用。這表明 Natterin 可能在七鰓鰻中發揮類似的功能。IL-27通過免疫信號傳導對代謝活性的調控機制為探索 Natterin 的代謝作用提供了相關框架。這表明免疫蛋白不僅介導免疫反應,還可能通過與脂肪細胞的相互作用來調節代謝過程。這些見解強調了七鰓鰻免疫系統與新陳代謝之間的相互作用,并支持 Natterin 作為代謝調節劑的潛在作用。
WAT 的褐變是指主要儲存脂肪的白色脂肪細胞轉化為代謝活躍并產生熱量的米色或棕色脂肪細胞。Natterin 通過靶向人體內一種關鍵的鐵轉運蛋白 TFR1 觸發鐵依賴性 WAT 褐變過程。TFR1 調節棕色和米色脂肪細胞中的鐵穩態、產熱和命運決定。這表明 Natterin 通過與 TFR1 結合來調節七鰓鰻的鐵代謝過程。WAT 褐變的激活會增加能量消耗,因為米色或棕色脂肪細胞的代謝更活躍,可以燃燒儲存的脂肪以產生熱量。能量消耗的增加可能有助于維持體重。這些發現凸顯了免疫蛋白的多功能性及其在調節代謝過程中的潛在參與,從而為制定解決代謝相關問題的策略提供了理論基礎。
最近的研究越來越強調免疫系統在調節代謝過程,特別是脂質代謝和能量穩態方面的關鍵作用。除了病原體防御之外,免疫細胞還通過分泌細胞因子和趨化因子影響代謝途徑。例如,IL-6、IL-12和IL-27等白細胞介素不僅是免疫反應的關鍵調節因子,還有助于脂肪組織重塑和能量消耗,特別是通過將 WAT 褐變為代謝活性的棕色或米色脂肪細胞。我們的研究表明 Natterin 在驅動 WAT 褐變方面具有潛在作用,并提出免疫蛋白可能通過與鐵代謝的關鍵調節因子 TFR1 的相互作用來影響脂質代謝。這種免疫代謝串擾可能代表了一種重要的進化適應,其中免疫信號通路有助于調節能量消耗,以支持環境適應,例如寒冷氣候下的體溫調節。需要進一步的研究來闡明這些免疫代謝相互作用背后的分子機制,并追蹤它們在物種之間的進化意義。這些見解可以增強我們對免疫系統在調節新陳代謝、適應性生理反應及其對全身能量平衡的更廣泛影響方面的多功能作用的理解。
總體而言,我們的研究結果提供了七鰓鰻器官的綜合細胞圖譜,為該物種的細胞組成和進化提供了見解。這項工作強調了七鰓鰻作為模式生物研究脊椎動物進化、免疫系統功能以及治療發現潛力的重要性。
參考文獻:
Pang Y, Qin Y, Du Z, Liu Q, Zhang J, Han K, Lu J, Yuan Z, Li J, Pan S, Dong X, Xu M, Wang D, Li S, Li Z, Chen Y, Zhao Z, Zhang Z, Chuan S, Song Y, Sun M, Jia X, Xia Z, Zhan L, Yue Z, Cui W, Wang J, Gu Y, Ni M, Yang H, Xu X, Liu X, Li Q, Fan G. Single-cell transcriptome atlas of lamprey exploring Natterin- induced white adipose tissue browning. Nat Commun. 2025 Jan 17;16(1):752. doi: 10.1038/s41467-025-56153-w. PMID: 39820434; PMCID: PMC11739602.




