CAR-T和TCR-T免疫療法識別腫瘤的機制及區別
文章來源公眾號:生物快評 作者:sw
CAR-T,Chimeric Antigen Receptor (CAR) T-Cell,全稱為嵌合抗原受體T細胞免疫療法,是一種革命性的癌癥治療方法,屬于細胞免疫治療和精準醫療的范疇。它被譽為是“活的藥物”。那么這個“嵌合抗原受體”怎么去理解,是按照“嵌合,抗原受體”去理解還是按照“嵌合抗原,受體”去理解呢?
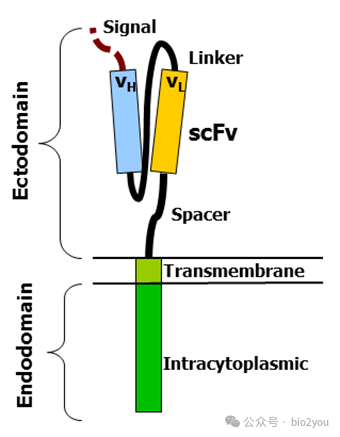
CAR-T的CAR的指的T細胞上的受體,從N端到C端分別是scFV,即結合抗原的部分,來源是抗體分子的結合部分,然后是鏈接端(spacer),最后是胞內的分子,所以我們可以發現這個CAR由3段蛋白結構域串聯而成,而不是“自然存在的蛋白分子”,這種人工“手搓”起來的分子叫做嵌合的抗原受體,簡稱“嵌合抗原受體”。
- T細胞識別腫瘤的背后邏輯
我們知道T細胞、B細胞是我們人體的重要的免疫細胞,T細胞是負責殺傷腫瘤、病毒感染的細胞等作用。
- T細胞受體,T-Cell Receptor (TCR)
那么T細胞怎么去識別腫瘤呢?很好理解T細胞依靠的自身的受體去識別,這種受體是叫做T-Cell Receptor (TCR)。同時我們知道腫瘤、細菌、病毒等分子在蛋白質上存在各種差異,并且這種差異的數目是“海量、天量的”,TCR是怎么識別“海量、天量的”外源、異源分子呢?
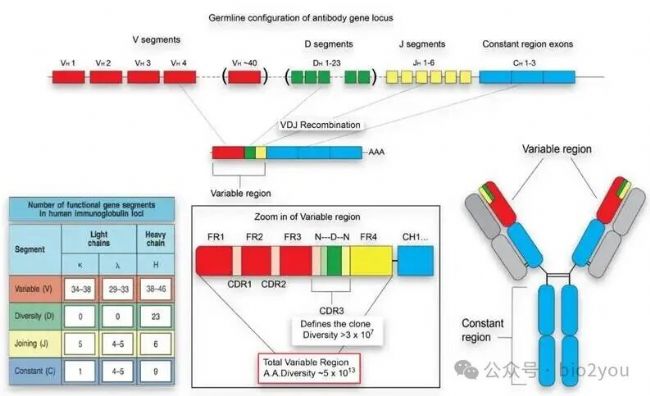
打敗魔法的必須也是魔法,“海量”的異源分子就需要用“天量”的TCR受體去識別。我們看下面這張圖:第一排位基因組上TCR的元件分子,有紅色的V區(可變區)、綠色的D區(多樣性區域)、黃色J區(鉸鏈區),藍色C(恒定區)。基因組上每個區域的元件都有好多種備份,那么每一個T細胞成熟過程中,都會在V、D、J中隨機抽取一個完成VDJ重排,然后C區位統一不變的部分。那么這種大自然“手搓”的分子叫做TCRα,另一條大自然“手搓”的類似分子叫做TCRβ。兩者在T細胞膜上組合為TCR受體(α、β)。
學過高中概率與統計中的古典概率學中的“排除組合問題”,我們可以知道TCRα的理論數目是V區元件數目乘以D區元件數目,再乘以J區元件數目,然后同理TCRβ。所以我們就可以得到天量的TCR受體,且每個T細胞上僅有一種唯一的TCRα/β。
- CD3-TCR復合物
剛才我們已經解決了多樣性的問題,那么單靠TCR分子結構,還無法完成胞外到胞內的信號傳導,還需要TCR和其他的復合物形成“TCR復合物”才可以完整發揮功能。這個復合物叫做“CD3-TCR復合物”。
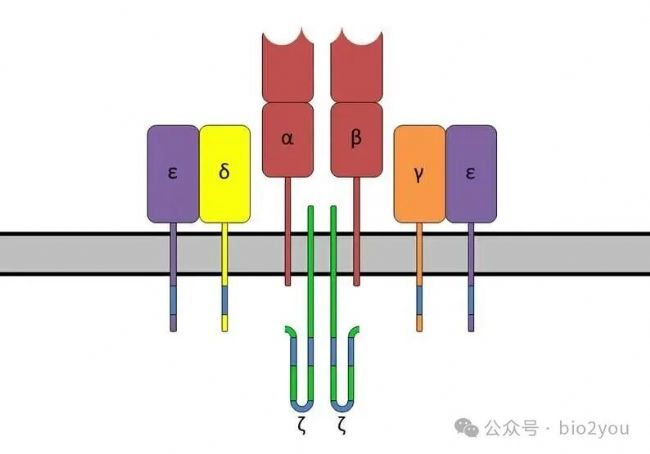
https://en.wikipedia.org/wiki/CD3_%28immunology%29
CD3-TCR復合物,中間的α、β是叫做TCRα、TCRβ,其余的γ、δ、ε和ζ分別是叫做CD3γ, CD3δ、CD3ε和CD3ζ。CD3的分子沒有多樣性。
那么CD3-TCR分別怎么識別腫瘤分子的呢?
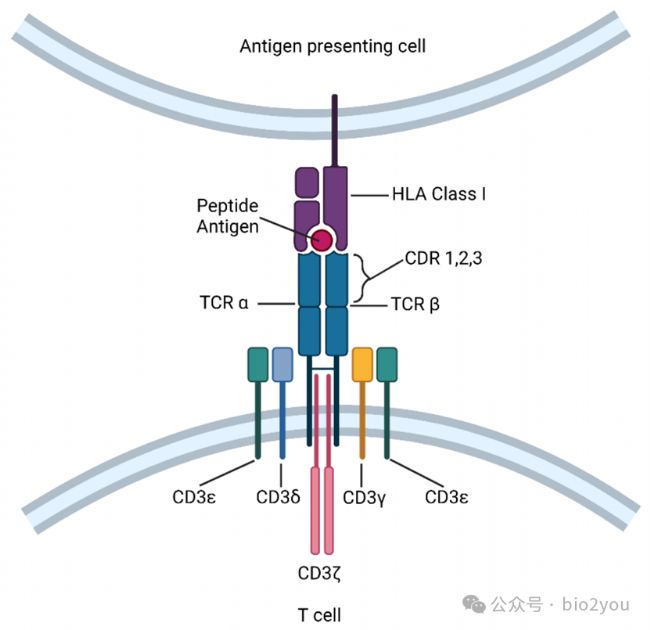
Cells 2021, 10(9), 2379; https://doi.org/10.3390/cells10092379
TCR識別的是其他細胞(抗原呈遞細胞)通過HLA-I分子呈遞的“腫瘤多肽“分子。那么這里面就有一個核心問題:腫瘤分子是什么,比如某個腫瘤細胞因為突變產生了一個突變的蛋白質,但是這個蛋白質并不會變化太多,可能僅僅某1-2個氨基酸和正常蛋白質不同,那么HLA-I怎么能夠呈遞這些分子呢?
- 人類白細胞抗原I類分子
HLA-I分子,The human leukocyte antigen (HLA) ,全稱為人類白細胞抗原I類分子,是存在于人體幾乎所有有核細胞表面的一種至關重要的蛋白質復合物。它是主要組織相容性復合體在人類身上的特定名稱,因此也常被稱為 MHC-I 類分子( (Major Histocompatibility Complex class I)。
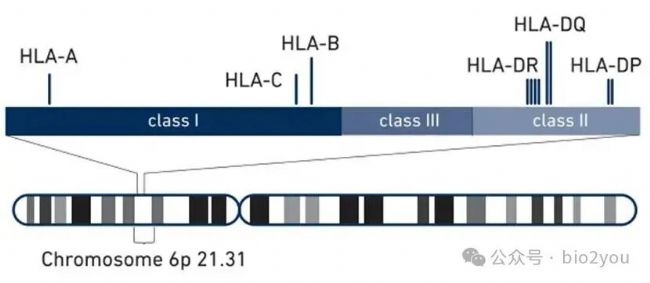
https://www.10xgenomics.com/blog/more-complete-hla-analysis
HLA-I分子共有三個經典的分子,即HLA-A, HLA-B, HLA-C,但是他們沒有什么多樣性,每個人的細胞上同時只有這三個分子。這三個HLA-A,B,或者C,均和B2M形成復合物。
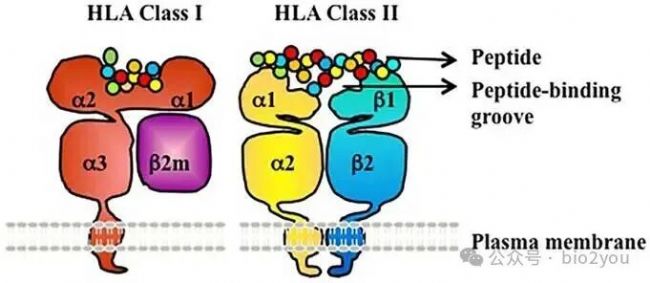
Frontiers in Immunology DOI:10.3389/fimmu.2016.00030
MHC-I分子,MHC-I和B2M形成MHC-I:B2M復合物。這里的槽子中間(α1、α2)會呈遞出“腫瘤多肽”。實際上上這個多肽不是僅僅腫瘤多肽,而是細胞中的一切可能得、可以被呈遞的多肽,如果呈遞的是自家蛋白質的部分肽,那么就忽視。如果是呈遞了非自己的多肽,那么T細胞就“激動”起來了。
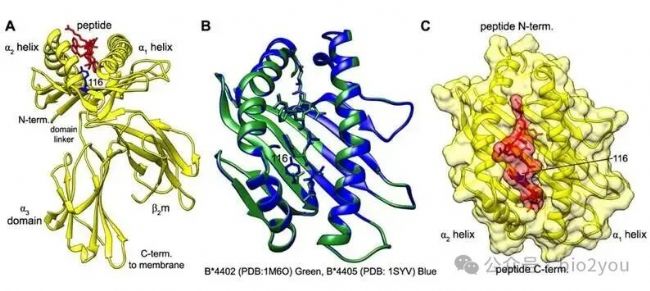
Bailey, A., Dalchau, N., Carter, R. et al. Selector function of MHC I molecules is determined by protein plasticity. Sci Rep 5, 14928 (2015). https://doi.org/10.1038/srep14928
HLA-I:B2M呈遞多肽依靠的也是物理作用,比如這里的HLA-I形成的槽子(有兩個α螺旋和一些β sheet形成),這個槽子只是一些滿足槽子結構的氨基酸可以牟定在這個槽子中。比如這里所示的紅色的氨基酸“躺在”黃色的槽子里。
- HLA-I呈遞抗原的背后之路:來吧!展示
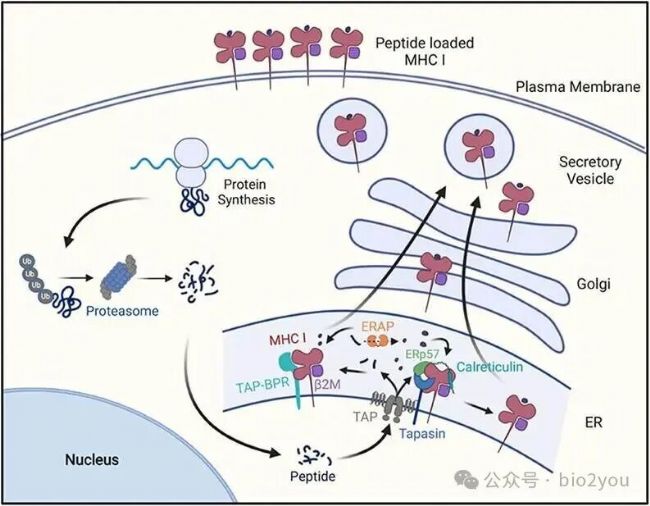
Front. Immunol.,doi.org/10.3389/fimmu.2021.636568
體內蛋白質經過泛素化降解途徑形成短肽,短肽隨機被運輸到ER內質網中,然后多肽在ERAP裁剪為統一長度,然后在ER中加載到HLA-I:B2M復合物中,最后多肽:HLA-I:B2M三元復合物經過高爾基體運輸到胞外,展示自己的內心,向T細胞表忠心,好像在說:你快過來看看,我還純粹嗎?”
T細胞拿著大棒子,來核查這些細胞是否都是呈遞自家的原件,如果不是,那么就要被T細胞消滅。
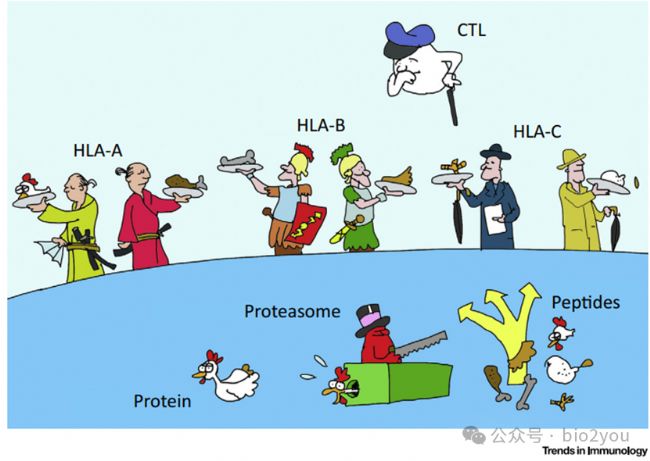
http://dx.doi.org/10.1016/j.it.2016.08.010
因為細胞上僅有3個HLA-I分子,即HLA-A、HLA-B和HLA-C。那么是否呈遞的多肽僅有3個呢?實際上HLA-A呈遞的分子是“簡并性”的,意思就是滿足一定條件的多肽,都會被呈遞,所以呈遞的多肽實際上是非常多的。但是是否還是天量的呢?可能不如TCR種類多。可能進化上夠用了,湊合著過吧,細胞就這樣處理了。
但是我們要認識到,雖然每個人僅有3個HLA-I經典分子,但是人群中HLA-A、B和HLA-C具有非常大的多樣性。一般來說,人體內的蛋白編碼基因都差不多功能,氨基酸序列類上非常相似,比如GAPDH蛋白在美國人、高加索人和藏區人民細胞中幾乎一模一樣。但是HLA-I就不是這樣邏輯了,比如你的HLA-A和我的HLA-A大概率是存在差異,那么我們呈遞的多肽可能就有差異,這個差異還不算小,甚至你和你媽的HLA-A都只有一半一模一樣。進化上這樣做的結果就是:我們群里中可以抵抗的疾病、細菌、病毒的概率增大了,如此高的多樣性確保了整個種群在面對不斷變異的病原體時,總有一部分個體的HLA能夠呈遞病原體的肽,從而啟動免疫應答,避免整個種群被一種病原體毀滅。通過“人類命運共同體”我們地球人功能抵抗外界環境。
這些HLA-A\B\C的序列到底有多少中呢?有興趣的可以到這里來看看IPD-IMGT/HLA(https://www.ebi.ac.uk/ipd/imgt/hla/。)
因為HLA-I在人群中的多樣性,導致了優勢就是我們地球人的抗風險能力增強了,壞處就是“我們不一樣了”,比如器官移植的時候,我們會互相排斥。移植排斥反應的本質就是受體的免疫系統識別了供體細胞的“外來”HLA,并發起攻擊。因此,移植前必須進行HLA配型,匹配度越高,移植成功率越高。
- TCR-T殺傷腫瘤細胞的原理
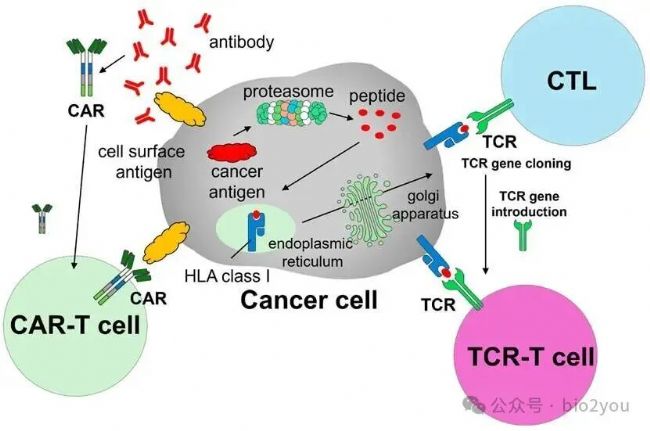
Cancers 2025, 17(12), 1945; https://doi.org/10.3390/cancers17121945
天然的T細胞可以識別腫瘤的叫做CTL(Cytotoxic T Lymphocyte),它是天然的TCR受體。如果這個CTL在腫瘤部位,那就是相對于這些CTL已經可以成功識別腫瘤了,這些CTL一般叫做TIL(Tumor-infiltrating lymphocytes (TILs))。
我們再強調了點:就是TCR識別HLA-I:B2M:peptide三元復合物,這個TCR是需要識別HLA-I:peptide的,意思就是TCR需要識別HLA-I也要識別peptide,且兩者是關聯的狀態下。
如果某些情況下,我們知道一個腫瘤發生的時候,它展現的多肽和對應的HLA-I分子,但是我們體內的T細胞缺少識別它的TCR。那么我們就可以抽出T細胞轉導一個TCR序列進入T細胞,重新武裝的T細胞表達了TCR,它就叫做TCR-T。經過上面的介紹,一般我們轉導TCRα,β兩個基因進入T細胞,然后把內源的TCRαβ敲除,目的是為了減少內源和外源兩套TCR分子錯配。因為TCR需要識別HLA-I:peptide復合物。如果我們鑒定到了一個腫瘤抗原和HLA-I的復合物關系,那么我們也找到了一個TCR能夠識別,在真實的產品中,我們需要對病人進行腫瘤抗原鑒定和對他們的HLA-I進行鑒定,如果兩個夠匹配,才可以使用轉染這個外源TCR,得到TCR-T。
比如這里6G4是一個鑒定出來的TCR的代號,它識別多肽LTQDLVQEKYLEY,這個多肽需要由HLA-A*01:01展示,這個多肽是MAGE-A1基因的部分多肽。這個MAGE-A1 是最早被發現的癌睪抗原之一。它是一個非常特殊的分子,該基因在成人除睪丸和胎盤以外的所有健康組織中完全沉默。由于睪丸和胎盤是免疫豁免器官,不表達MHC I類分子,因此細胞毒性T細胞無法接觸到這些抗原。在腫瘤中:這些基因被異常激活,在多種類型的癌細胞中高表達。因此,MAGE-A1可以被免疫系統視為一個 “非己” 的信號,成為一個理想的免疫攻擊靶點。
一般情況下TCR-T很難做出通用性的TCR-T。背后的原因在外源和內源的TCR會錯配,那么就要把內源TCR敲除(避免異體的天然T細胞對患者細胞攻擊),然后轉導外源的TCR(用于識別腫瘤抗原)。但是我們無法100% 敲除內源TCR,所以總有一點點TCR殘留,那么殘留的TCR和外源的TCR都會和CD3形成復合物,就算通過CD3分選也難以成功,而殘留的TCR-CD3復合物很少比例下都會產生排斥作用。
所以為了克服這個問題,也有一些公司開始開放TCR蛋白,讓TCR蛋白在體內結構在T上形成瞬時的TCR-T。
- CAR-T繞開了HLA-I的限制
因為HLA-I必須和多肽形成復合物,且有配對關系,TCR才能識別。CAR-T就是將抗體的識別區域替換了TCR的序列。然后將CD3的核心分子CD3zeta放在胞內端,得到CAR-T。
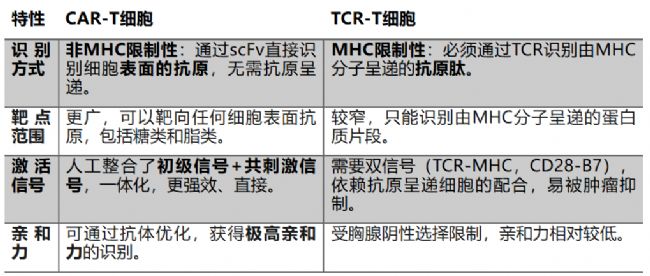
CAR-T與TCR-T細胞殺傷的區別