
果蠅單細胞三維時空多組學圖譜揭示細胞類型分化的全景關鍵調節因子

期刊:Cell
影響因子:45.5
主要技術:Stereo-seq、scRNA-seq、scATAC-seq
導語
多細胞生物的發育是一個高度復雜的過程,受到無數基因和通路在空間和時間上的嚴格調控。單細胞水平上基因表達的空間環境對于理解其生物學相關性至關重要,但在標準的單細胞測序過程中往往丟失。以前,我們利用了空間增強分辨率組學測序(Stereo-seq),一種基于測序和模式化 DNA 納米球(DNB)陣列的空間轉錄組學平臺,具有高空間分辨率和靈敏度,來解決這個問題。在這里,我們將對果蠅時空轉錄組的研究擴展到覆蓋其從胚胎到蛹期的整個發育過程。我們使用 Stereo-seq 和 Spateo,這是一個用于分析單細胞多模態數據的計算流程,重建了單細胞空間分辨率的3D轉錄組。我們進一步將單細胞 Stereo-seq(scStereo-seq)數據與單細胞 RNA 測序(scRNA-seq)和單細胞轉座酶可及染色質測序(scATAC-seq)數據相結合,創建了一個果蠅胚胎發生的多組學圖譜。這個圖譜包括超高通量空間背景下轉錄組和表觀遺傳信息,這使我們能夠建立多組學細胞狀態軌跡,并具有相關分子調控網絡的時空動態。
為了從 3D 時空多組學的角度闡明各種細胞類型的發育調控,我們將中腸作為研究的模型。果蠅中腸通過不同類型的細胞和它們形成的區域發揮著多種功能,例如吸收營養的腸上皮細胞(ECs)、感知刺激和分泌激素的腸內分泌細胞(EEs)以及與金屬離子穩態和胃酸分泌相關的銅細胞。在變態過程中,幼蟲中腸細胞經歷自噬依賴性細胞死亡,成年中腸細胞則從一群專門指定的干細胞中重新構成,這些干細胞被稱為成年中腸祖細胞(AMPs)。然而,分化的時機和調控機制尚未完全闡明。我們重點關注發育中的中腸,從多組學角度研究了其細胞類型多樣化、基因調控網絡(GRNs)和區域化。通過多模態分析,我們確定了參與中腸發育細胞類型特異性調控的多個潛在因素,并通過突變分析驗證了同源框(HD)轉錄因子(TF)exex 是銅細胞的關鍵調節因子。這個廣泛的圖譜提供了一個豐富的資源,并作為系統平臺,用于研究具有超高通量時空分辨率的集成單細胞數據的果蠅發育。
主要技術
Stereo-seq、scRNA-seq、scATAC-seq
研究結果
1. 以單細胞分辨率重建從胚胎發育到變態階段的果蠅三維空間轉錄組
為了構建果蠅發育的多組學圖譜,我們首先通過收集發育階段不同的樣本,包括胚胎(0.5 到 2 小時間隔)、幼蟲(L1 到 L3 的早期和晚期階段)和蛹(P12 到 P72),來擴展和增強 3D 空間轉錄組(圖 1A)。為了實現單細胞空間分辨率,我們對每個芯片進行了核染色和細胞分割,從而允許進行更精確的單細胞轉錄組分析。利用 Stereo-seq 平臺,我們生成了 43 個胚胎、9 個幼蟲和 5 個蛹樣品的全器官單細胞空間轉錄組,總共有 3812505 個細胞單元(圖 1A)。然后,我們將來自單個樣本所有切片的細胞單元組合起來,基于基因表達譜和空間位置進行無監督聚類,并手動注釋聚類。通過將空間位置和細胞注釋與 Spateo 結合,我們實現了 3D 空間轉錄組來重建具有精細解剖形態的各種組織(圖 1A)。基于這個全面的時空轉錄組數據集,我們生成了一個在已建立的 in situ 數據庫中未報道空間表達模式的 338 個基因列表,并在 3D 中重建了它們的模式。因此,我們為果蠅樣品生成了 3D 空間轉錄組,涵蓋了從胚胎到蛹的整個發育階段。
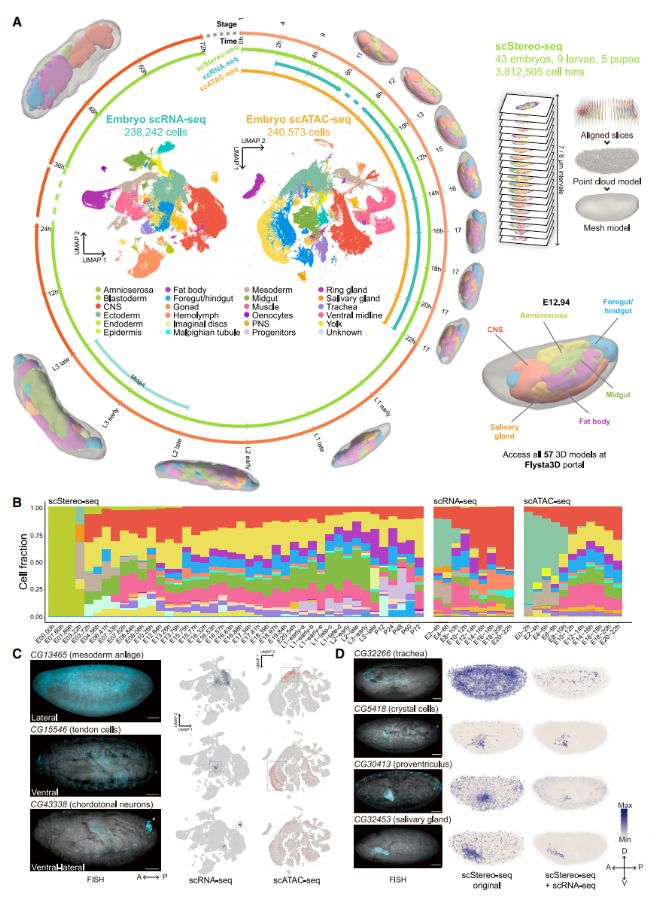
圖 1
2. 果蠅胚胎發育的單細胞時空多組學圖譜
為了用更深入的轉錄組和表觀基因組信息來增強我們的單細胞三維空間轉錄組數據。我們在胚胎發生過程中,每隔2小時收集胚胎樣本,并對其進行了基于滴液的scRNA-seq和scATAC-seq 測序(圖 1A)。經過質量控制后,我們獲得了238242個單細胞轉錄組,其中scRNA-seq的中位數為每個細胞6841個獨特的分子標識符(UMIs)和 1707 個基因。我們還獲得了240573個單細胞染色質可及性配置文件,其中 scATAC-seq 的中位數為每個細胞11772個片段。為了在幼蟲階段進行更全面的分析,我們從 NP1-Gal4 > UAS-mCherry-shRNA(其中 NP1-Gal4 是中腸 EC 特異性驅動器;shRNA,短發夾 RNA)的 L3 中腸中生成額外的控制 scRNA-seq 數據集。經過質量控制措施,我們獲得了 17988 個單細胞轉錄組,其中中位數為每個細胞8846個UMIs和1795個基因。
在胚胎發生過程中收集的聚合 scRNA-seq 數據,我們首先進行了粗略的無監督聚類,并在 UMAP 圖中生成了 45 個細胞簇。我們對這些簇進行了注釋,并在三個級別(細胞類型-組織-胚層)對注釋進行了分類(例如,胃盲囊-中腸-內胚層)(圖 1A)。我們還對聚合scATAC-seq數據進行了粗略的無監督聚類。類似地,我們在 UMAP 圖中產生了40個不同注釋的簇(圖 1A)。我們收集的數據涵蓋了主要組織,這從每個組織所代表細胞的比例及其在發育階段的變化中可以看出(圖 1B)。
我們進一步通過亞聚類分析了組織細胞類型異質性。在scRNA-seq數據中,我們能夠廣泛地描述胚胎組織的細胞類型組成,包括稀有細胞類型。為了驗證我們識別的亞簇,我們編制了一個在兩個數據集中都識別出的常見細胞類型標記基因列表,并使用熒光原位雜交(FISH)驗證了 3 個之前未報道的細胞類型標記基因的表達特異性(圖 1C)。總之,我們生成了果蠅胚胎發生過程中 scStereo-seq、scRNA-seq 和 scATAC-seq 數據集的綜合圖譜。我們多組學數據的高粒度和時間連續性為細胞類型和發育年齡依賴的這些多組學數據整合打開了可能性。
3.多組學數據集成與構建組織分化軌跡
為了整合 scStereo-seq 和 scRNA-seq 數據,我們使用 NovoSpaRc 選擇推斷發育年齡差異為 1 小時的細胞進行整合。為了評估這種整合,我們選擇了 9 個之前未表征的組織特異性基因,并使用 FISH 建立了它們表達模式的基準。我們發現,與僅使用 scStereo-seq 數據相比,整合數據明顯減少了信號背景,增強了組織富集,并改善了空間模式,使其與 FISH 結果更加相似(圖 1D )。
為了整合 scRNA-seq 和 scATAC-seq 數據,我們將 scATAC-seq 峰矩陣中的表達矩陣進行插補,并將其與 scRNA-seq 表達矩陣共嵌入同一UMAP 空間進行聚類(圖 2A)。接下來,我們旨在將 scRNA-seq 和 scATAC-seq 數據中每個胚層的所有胚胎組織細胞狀態組織成連續的發育軌跡。我們將 PhyloVelo29 應用于整合數據,為三個胚層建立了速度矢量場,并根據標記基因重新注釋細胞簇以及它們沿速度軌跡的順序。有了這些速度矢量,我們能夠確定每個組織的分化軌跡,并將細胞類型按時間順序排列(圖 2B)。由于細胞類型分化的復雜性,細胞類型分化分支在 3D UMAP 空間中可視化效果更好。這些結果表明,我們的數據集可以作為探索細胞信號網絡的系統框架。
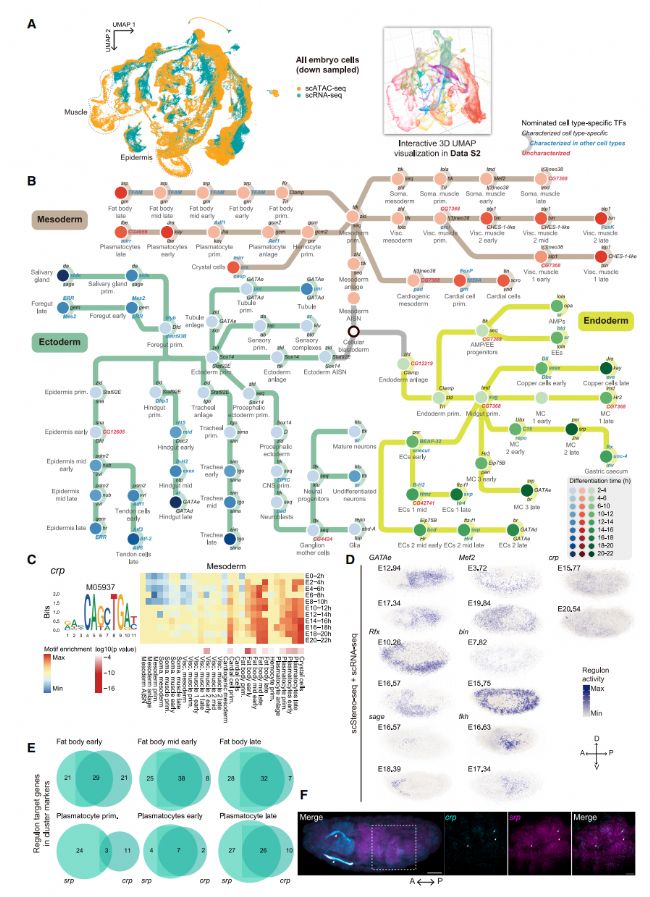
圖 2
4. 組織分化過程中細胞類型特異性的轉錄因子活性
為了解析沿組織分化軌跡由 TFs 指導的細胞類型特異性調控活動,我們仔細審查了我們的整合 scRNA-seq/scATAC-seq 數據。我們首先在 scATAC-seq 數據中識別了表現出細胞類型特異性活動的 TF 結合基序。我們進一步通過 scRNA-seq 數據中細胞類型標記基因的靶基因富集過濾候選 TF 列表。這種整合分析中重疊的候選 TF 代表了沿分化路徑最活躍的細胞類型特異性 TF,包括既定的調節因子和尚未表征的潛在因子(圖 2B)。
代表性的細胞類型特異性 TF 活動包括 GATAe 在 Malpighian 腎管中的基序、Rfx 在外胚層 PNS 和 CNS 中的基序和 sage 在唾液腺中的基序。在中胚層中,我們發現了 Mef2 在軀體肌肉中的基序富集、bin在內臟肌肉中的基序和 srp 在脂肪體和血細胞中的基序。內胚層表現出 GATA 家族 TFs grn、fkh 和 GATAe 的基序富集,這些 TF 調節晚期內胚層分化。我們還發現了幾個之前未表征的 TF,它們在胚胎發生過程中具有潛在的時空特異性功能。TF crp 在多個組織中廣泛表達,并已知在氣管管道中指定終細胞,在中胚層脂肪體和血細胞中表現出潛在的調控功能(圖 2C)。
為了進一步探索這些 TF 的空間調控子活動,我們將 SCENIC45 應用于整合 scStereo-seq/scRNA-seq 數據,揭示了 TF 調控子活性的空間模式與組織特異性基序富集分析一致(圖 2D)。這些不太確定的 TF 的空間表達模式也通過伯克利果蠅基因組計劃(BDGP)原位數據庫進行了探查,所有這些 TF 在其推斷功能的階段都表現出弱信號或普遍表達模式。因此,我們的多組學數據為闡明這些 TF 的組織特異性調控作用提供了額外的證據。
隨后,我們將 Pando47應用于整合 scRNA-seq/scATAC-seq 數據,深入探討了識別的 TF 的詳細調控子。在脂肪體和血細胞特異性調控子活動中,我們發現 crp 和 srp 位于同一 GRN 中。值得注意的是,我們發現 srp 和 crp 的調控子中的靶基因在早期脂肪體中 largely 相互重疊,并且這種重疊在中胚層組織(包括脂肪體和血細胞)的發育軌跡中增加(圖 2E)。我們識別的 srp 的調控子與從早期脂肪體發育開始誘導脂肪細胞形成的作用一致,crp已知影響細胞生長和組織大小控制。我們的分析暗示 crp 和 srp 在脂肪體和血細胞發育過程中在同一個 GRN 中的作用越來越協調。與這一發現一致,FISH 結果證實了 crp 和 srp 表達細胞在晚期胚胎中的重疊(圖 2F)。通過追蹤組織分化軌跡,我們確定了在細胞類型分化過程中既定的和潛在的 TF,揭示了它們的組織特異性和協調的調控網絡。
5. 通過時空細胞類型圖譜研究組織分化的起源和模式
我們選擇了脂肪體(哺乳動物肝臟的對應物)和前腸/后腸(哺乳動物胃/大腸的對應物,兩者均為外胚層來源作為模型,并將它們的細胞類型與胚胎 scStereo-seq 樣本進行對齊(圖 3A 和 3B)。在各個發育階段對組織細胞類型進行映射為在時空背景下檢查分化起源的分布提供了獨特的機遇。具體來說,具有集中分化起源(如干細胞)的組織將在發育的不同階段產生空間上不同的細胞類型。相比之下,具有分散分化起源的組織將在不同階段產生細胞類型的混合物(圖 3C)。
以前的研究確定,脂肪體細胞起源于在整個組織中延伸的重復片段中的祖細胞。這表明脂肪體分化是分散的。為了驗證這一點,我們使用鄰域富集分析量化了脂肪體細胞類型的空間聚集水平,其中較高的分數表示更高的空間聚集水平。我們發現脂肪體的鄰域富集水平較低。一致地,我們在 3D 模型中發現,脂肪體細胞類型的各個階段分布分散且混合(圖 3A)。這進一步得到了 scStereo-seq 數據中細胞單元 CytoTRACE 分數的空間分布的支持。CytoTRACE 利用可檢測到的基因表達數量作為分化潛力的穩健指標。我們觀察到脂肪體 scStereo-seq 數據中具有不同分化潛力的細胞相互混合(圖 3D)。
另一方面,胚胎前腸/后腸分化起源的空間分布尚未完全繪制。以前的研究確定了成年腸道中消化道干細胞的生態位,其中在空間上定義的干細胞群分別產生成年前腸和后腸。也有報道稱,胚胎后腸來自受多種信號通路調節的狹窄環狀區域,這些通路控制后腸節段的生長。胚胎器官發生過程中腸道干細胞的數量和分化起源的位置仍然不清楚。我們的數據表明,前腸/后腸細胞類型具有顯著更高的鄰域富集。不同發育階段和分化潛能的細胞類型也表現出不同的空間分布(圖 3A 和 3D)。這些發現表明,胚胎前腸/后腸以集中方式分化,為胚胎前腸和后腸中存在成簇而不是分散的分化起源提供了證據。總之,組織細胞類型的空間映射為分析分化起源和識別潛在干細胞生態位提供了直觀的視角。
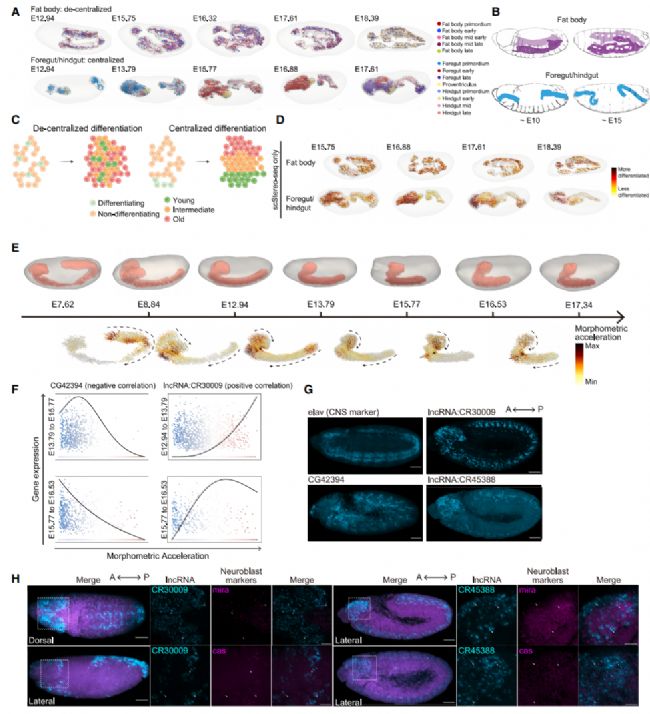
圖 3
6. 中樞神經系統形態測量學中的轉錄組動力學變化
我們在 7 個 scStereo-seq 樣本中對中樞神經系統進行了形態學分析,這些樣本的發育年齡跨度從 7 小時到 18 小時。在整個中樞神經系統發育過程中,我們觀察到最高加速區域從 VNC 的后端(E8.84 之前)轉移到大腦的前端(E13.79 之后)(圖 3E)。VNC 中加速和曲率分數的下降可能與胚帶收縮的完成有關,這表明早期發育過程中 VNC 的縮短主要依賴于后部細胞向前端遷移。相反,大腦前端區域加速和曲率分數的增加可能反映了晚期胚胎發生過程中大腦葉的細胞組織活動(圖 3E)。正如預期中的中樞神經系統形態學一樣,最高曲率和曲率分數的區域集中在 VNC 和大腦之間的彎曲關節處。形態學分析產生了一組與中樞神經系統形態計量動力學相關的時空表達變化基因。GO富集分析顯示,與中樞神經系統形態計量變化相關的基因在細胞命運決定和模式形成中高度富集。值得注意的是,基因表達術語在早期階段更富集,而信號轉導術語在后期階段更富集。這些觀察結果表明,中樞神經系統形態發生過程與內在細胞命運決定密切相關。這得到了已知中樞神經系統發育調節因子(例如,mira、tll 和 toy)對中樞神經系統形態計量分數的貢獻的進一步支持。此外,我們識別了多個未表征的因子。例如,編碼未表征跨膜蛋白的 CG42394 的表達水平與加速呈負相關,而 lncRNA:CR30009 的表達水平與加速呈正相關(圖 3F)。我們使用 FISH 驗證了這些潛在調節因子在中樞神經系統中的特異性表達,并觀察到 CG42394 表現出分段表達模式,而 lncRNA:CR30009 在中樞神經系統中更普遍地表達(圖 3G)。值得注意的是,潛在形態計量調節因子列表中除了 lncRNA:CR30009(之前報道在膠質細胞中富集并共定位與膠質細胞標記基因 repo)之外,還包括多個長非編碼 RNA(lncRNA)基因。在我們 scRNA-seq 數據中檢查這些 lncRNA 基因時,我們觀察到 lncRNA:CR30009 和 lncRNA:CR45388 的表達與神經母細胞和膠質母細胞標記基因的相關性最高。我們進一步使用 FISH 確認了它們與神經母細胞標記基因 mira 和 cas 的共定位(圖 3H)。這些觀察結果表明,這兩個 lncRNA 基因可能通過調節神經母細胞來影響中樞神經系統細胞遷移。
因此,對中樞神經系統的形態計量分析提供了一個獨特的視角來研究與細胞遷移相關的基因,并確定了已知和潛在的中樞神經系統細胞遷移調節因子。
7. 胚胎中腸細胞亞群的基因表達與空間布局
果蠅中腸是研究細胞分化調控機制廣泛使用的模型。在胚胎中腸發育過程中,細胞類型的解剖形態和空間分布高度動態(圖 4A)。在各種中腸細胞類型中,內分泌細胞(EEs)和腸上皮細胞(ECs)可以根據它們的標記基因和在成體中的生理功能進一步分類為多個亞類。然而,它們在胚胎和幼蟲發育過程中的動態變化仍然不清楚。在這里,我們首先旨在將胚胎中腸細胞類型分類為亞類,并使用 Dynamo 描繪它們的分化動態。我們集中在中腸 scRNA-seq 數據上進行高分辨率亞聚類、注釋和細胞狀態譜系描述(圖 4B)。這揭示了發育中中腸的 23 個細胞亞簇,并確定了 AMPs(由 esg 標記)、EEs(由 pros 標記并由特定內分泌基因表達區分)和 ECs(由消化酶和代謝相關基因標記和區分)的存在和分化路徑(圖 4C)。
已知一些果蠅中腸細胞類型占據不同的空間位置以執行其功能。例如,銅細胞專門位于中腸中部的小段中。為了探索中腸細胞亞簇的空間模式,我們通過從 scRNA-seq 到胚胎 scStereo-seq 數據的標簽轉移,將上述識別的亞簇映射到它們各自的空間位置(圖 4D)。正如預期的那樣,胃盲囊和銅細胞表現出高度的空間聚集(圖 4D)。在標簽轉移的 scStereo-seq 3D 模型中,我們觀察到了整個胚胎發生過程中細胞類型比例的動態變化,反映了這些細胞類型出現的時間不同。例如,EC(Try29F+)在大約 13 小時的發展中出現,而 EC(Acbp3+)直到大約 17 小時才形成(圖 4E)。鄰域富集分析表明,所有 EC 亞簇都表現出更高的聚集空間分布。在胚胎 scStereo-seq 樣本中,我們觀察到 Jon99Cii 和 Try29F 標記的 ECs 占據不同的空間位置,這表明這些負責產生消化酶的 ECs 傾向于占據中腸道的更后部位置(圖 4F)。我們進一步在 BDGP 原位數據庫中檢查了具有高鄰域富集分數的細胞亞簇標記基因的空間分布。
總結來說,我們對胚胎中腸中存在的各種細胞亞簇進行了分類和檢查。我們的分析揭示了占據不同空間位置的 EC 亞簇,并追蹤了它們在發育過程中的分布。
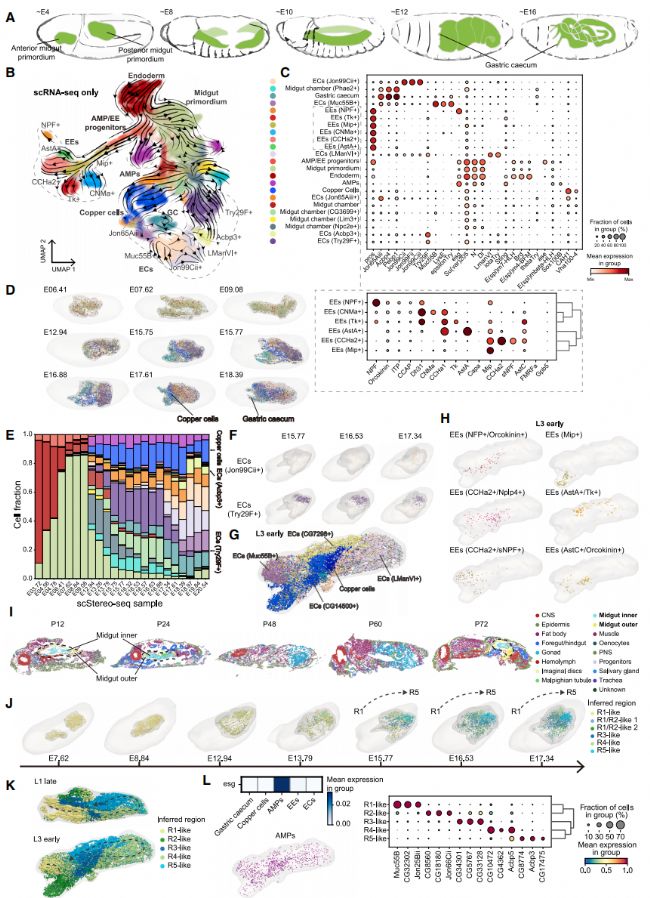
圖4
8. 幼蟲和蛹中腸細胞亞群的基因表達和空間布局
與胚胎相比,幼蟲中腸的大小顯著增長,并且在發育過程中顯示出更多樣化的腸細胞類型(圖 4G )。值得注意的是,不同的 ECs 沿著中腸的前后軸緊密聚集,正如它們分布的 3D 模型中所觀察到的。在成年中腸中,已經確定 EEs 可以被分類為幾個亞簇,每個亞簇表達獨特的肽激素組合,并占據中腸長度上的不同區域。類似地,在我們的幼蟲 scStereo-seq 數據中,EEs 也可以被分類為表達各種肽激素的亞簇。這些 EE 亞簇也顯示了不同的空間分布,類似于在成年中腸中觀察到的(圖 4H)。這些觀察結果表明,EEs 的空間模式可能在幼蟲中也會發生,類似于成年。
我們的蛹 scStereo-seq 樣本的空間轉錄組數據為研究中腸變態的調控提供了寶貴的資源。檢查蛹 scStereo-seq 樣本,我們觀察到蛹中腸分層為兩層,我們在手動注釋中將這兩層標注為“中腸內層”和“中腸外層”(圖 4I)。這些結構在 P24 階段后退縮,并在 P72 階段重新出現。之前的研究發現,在變態過程中,部分 AMPs 定位于圍繞幼蟲中腸的上皮層,該層退化成一種稱為黃色體的結構。蛹 scStereo-seq 樣本中觀察到的中腸結構可能反映了這種分層。檢查這兩層的表達譜,我們觀察到中腸內層群富集了金屬硫蛋白和鐵蛋白家族基因,而中腸外層群表達了多種抗菌通路基因(例如,DptA 和 Drsl2)。這些結果為中腸變態過程中基因調控的研究提供了線索。
總之,使用標簽轉移輔助的空間映射和注釋揭示了空間上不同的幼蟲和蛹中腸細胞類型及其在發育過程中的動態變化。
9. 中腸功能區的出現與空間分布
同時,區域的空間分布開始在同一時間點結晶,反映了成年中腸中觀察到的空間順序(從前到后為 R1 到 R5)(圖 4J)。這表明晚期胚胎中腸表現出與其成年對應物相似的區域分隔。我們檢查了已識別的胚胎中腸區域的標記基因的GO富集。例如,R1/R2 類區域在功能上富集于脂肪酸代謝;R3 類區域在功能上富集于離子運輸和 pH 調節,這與該區域的酸性性質一致;R5 類區域在功能上富集于金屬離子穩態。這些功能與成年中腸區域中的對應功能非常吻合。我們以類似的方式對幼蟲中腸區域進行了分析。成功識別了對應于 R1 到 R5 區域的模塊,這些模塊沿幼蟲中腸的前后軸依次排列(圖 4K)。在我們的 scStereo-seq 數據中,我們注意到雖然 AMPs(由 esg 表達標記)分布在整個中腸中沒有空間聚集(圖 4L),但位于每個已識別區域的 AMPs 表現出不同的表達譜(圖 4L)。這些發現表明,中腸干細胞的區域特異性發生在幼蟲階段。
總之,我們的 scStereo-seq 數據展示了胚胎和幼蟲中腸中的不同區域及其基因表達譜,這些譜決定了與成年人相似的局部亞器官功能。
10. 通過多模態分析鑒定exex作為銅細胞調節因子
為了尋找中腸發育過程中以前未表征的細胞類型特異性調節因子,我們追蹤了整合的 scRNA-seq/scATAC-seq 數據的內胚層軌跡(圖 5A )。我們首先在每個譜系內進行差異基序分析,以識別表現出細胞譜系特異性活性的 TFs。在早期中腸中,我們識別了多個與先前報道的調節 AMP 和 EE 分化的 TFs 相對應的基序,包括 ttk73 和 lola。在晚期中腸分化過程中,我們還識別了已知的中腸發育調節 TFs,如 fkh75 和 cad76。
檢查中腸譜系時,我們注意到銅細胞從早期中腸原基分化為高度特化的譜系(圖 5A )。我們觀察到分化后的銅細胞在胚胎發生約 10 小時出現(圖 4E)。這與從 BDGP 原位數據庫推斷的時間點一致,其中銅細胞標記 Vha100-4 在胚胎發育第 13 階段(約 10 小時)后出現。為了進一步追蹤銅細胞譜系回溯到中腸原基的起源,我們對中腸原基簇進行了亞聚類,產生了 7 個細胞群,每個細胞群沿著分化軌跡注定發展為各種中腸細胞類型(圖 5B)。其中,亞簇 2 導致了銅細胞的發展,我們觀察到特異性轉錄表達和 lab 基序活性的提高。這些發現與先前的發現一致,即同源盒(Hox)TF lab 是果蠅中腸中銅細胞的一個已確立的特異性決定因子,對于銅細胞的特化和維持既必要又充分。為了評估這一假設,我們檢查了沿著譜系中與銅細胞差異基序相對應的 HD TFs 的表達水平。其中,我們注意到 exex 在譜系起源處表現出顯著的高表達水平,在銅細胞分化后表達水平下降(圖 5C 和 5D)。這些觀察進一步暗示 exex 在胚胎到幼蟲階段的銅細胞發育中發揮作用。
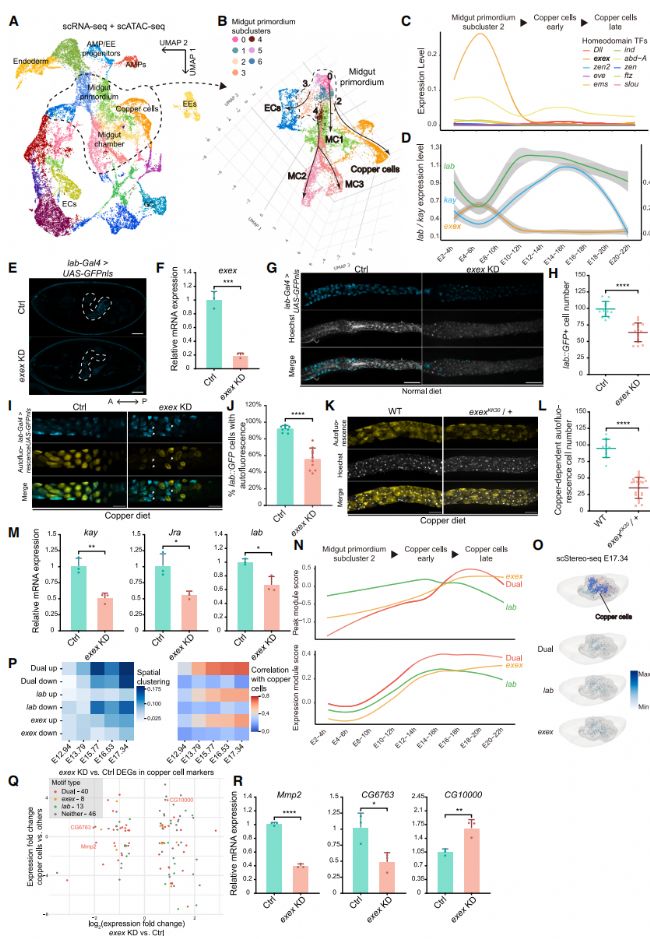
圖5
11. exex 通過調節 kay 和 lab 的表達來維持對銅細胞的特化和穩態
為了驗證 exex 的功能,我們使用 lab-Gal4 驅動 UAS-exex-shRNA 和 UAS-GFPnls 報告基因的表達,進行了銅細胞特異性敲低(KD)。檢查晚期胚胎,我們發現與對照胚胎相比,exex KD 胚胎在銅細胞區域的 lab::GFP 信號較弱且較少(圖 5E)。一致地,我們觀察到 RNAi KD 導致 L3 中腸中 exex 表達和 lab::GFP 陽性細胞數量顯著減少(圖 5F-5H)。剩余的 lab::GFP 陽性細胞與對照中觀察到的典型大、扁平、球形銅細胞特征相比,表現出明顯較小的尺寸和異常形態(圖 5G)。
然后,我們檢查了 exex KD 是否影響了銅細胞的生理功能。銅細胞在飲食銅處理后,當被紫外線激發時,會發出橙色自體熒光。這被認為是金屬硫蛋白與銅細胞中積累的銅離子相互作用的結果。在用銅飲食處理的 exex KD L3 中,我們觀察到 lab::GFP 陽性細胞數量和同時表現出橙色熒光的 lab::GFP 陽性細胞比例顯著減少(圖 5I 和 5J)。這表明在 exex 缺乏時,銅細胞中的銅離子穩態喪失。我們進一步研究了在攜帶 exexKK30(exex 功能喪失等位基因)的銅飲食 L3 中銅細胞的表型。這種品系在我們手中是純合致死的。在雜合 L3 中,我們注意到銅依賴性自體熒光細胞的顯著丟失和形態改變(圖 5K 和 5L)。總之,這些發現證實了 exex 在銅細胞發育中的關鍵作用。
隨后,我們探索了 exex 調節銅細胞特化的潛在機制。我們注意到 kay 和 Jra 的基序活性在銅細胞中排名靠前。這兩種 TFs 是已知的中腸中 lab 表達的激活因子。考慮到 exex 表達水平與 kay 和 lab 之間的時間不同步(圖 5D),exex 活性可能先于其他兩個在原初銅細胞中起作用。這一時間順序得到了在 kay 的 TSS 區域識別 exex 基序的支持。此外,exex KD 導致 kay 表達水平顯著降低。盡管在 Jra 的 TSS 區域沒有發現 exex 基序,但我們仍然注意到相似水平的表達減少。與這些發現一致,我們觀察到 lab 表達的中度但顯著的減少(圖 5M)。
綜上所述,這些觀察強調了 exex 在調節果蠅中腸中銅細胞發育和銅穩態中的關鍵作用,通過調節 kay 和 lab 的表達水平。
12. exex 和 lab 在銅細胞發育過程中的潛在調控
盡管 exex 和 lab 的表達水平顯示了時間序列,但它們的結合基序在銅細胞譜系中持續活躍。因此,我們探索了 exex 是否也和 lab 協同發揮調控功能。為了檢驗這一假設,我們檢查了所有包含 lab 或 exex 基序的峰,發現同時包含這兩種 TF 基序的峰(雙重基序)與銅細胞標記基因的重疊程度比僅包含 lab 或 exex 基序的峰(單一基序)更大。與這一觀察結果一致,由雙重基序調節的銅細胞標記基因表現出比僅由 lab 基序調節的基因更高的染色質可及性和表達水平(圖 5N)。
此外,我們使用 scStereo-seq 數據可視化了在銅細胞中由雙重或單一基序上調或下調的標記基因的空間表達模式。我們觀察到,具有雙重基序的上調基因在發育過程中表現出空間聚集趨勢的增加,而具有單一基序的基因沒有表現出空間聚集模式(圖 5O 和 5P)。與具有單一基序的基因相比,具有雙重基序的基因在時間上與銅細胞的空間相關性逐漸增加和增強(圖 5P)。這些發現表明,由雙重基序調節的基因在發育過程中對銅細胞的特異性逐漸增強。
為了進一步檢測 exex 銅細胞特異性 KD 導致的全球變化,我們對銅細胞進行了批量 RNA-seq。在 exex KD 和對照組之間檢測到的 415 個差異表達基因(DEGs)中,我們觀察到有 107 個(25.8%)與銅細胞標記基因重疊。在這些基因中,我們注意到具有雙重基序的失調基因數量比具有單一基序的基因更多(圖 5Q)。檢查代表性的雙重基序基因,包括金屬內肽酶基因 Mmp2 和 CG6763,以及一個未表征的糖基轉移酶基因 CG10000,我們注意到在 exex KD 上的顯著表達失調(圖 5Q),這得到了 qPCR 的證實(圖 5R)。我們進一步觀察到這些基因的銅細胞差異可及性(DA)峰中存在雙重基序。這些觀察結果表明,除了調節 lab 表達外,exex 的功能可能還包括在銅細胞發育過程中作為 lab 的轉錄共調節因子。
總之,通過體內功能驗證,我們確認了 exex 在銅細胞發育調控中的關鍵作用。這些結果強調了我們的多組學數據在識別發育組織中的細胞類型水平調節因子方面的實用性。
導語
多細胞生物的發育是一個高度復雜的過程,它受到無數基因和通路在空間和時間上的嚴格調控。在這里,我們介紹了 Flysta3D-v2,一個涵蓋模式生物果蠅從胚胎到蛹期發育全過程的全面多組學圖譜。我們的數據集包括 3D 單細胞空間轉錄組、單細胞轉錄組和單細胞染色質可及性信息。通過整合多模態數據,我們生成了整個有機體發展中連續的虛擬 3D 模型。我們進一步構建了組織發育軌跡,揭示了細胞類型分化的詳細特征。重點關注中腸,我們使用多組學數據分析確定了參與中腸細胞類型調控的轉錄因子,并通過突變研究驗證了 exex 是銅細胞發育的關鍵調節因子。這個廣泛的圖譜提供了一個豐富的資源,并作為系統平臺,用于研究具有超高通量時空分辨率的集成單細胞數據的果蠅發育。
參考文獻:
Wang M, Hu Q, Tu Z, Kong L, Yu T, Jia Z, Wang Y, Yao J, Xiang R, Chen Z, Zhao Y, Zhou Y, Ye Q, Ouyang K, Wang X, Bai Y, Yang Z, Wang H, Wang Y, Jiang H, Yang T, Chen J, Huang Y, Yin N, Mo W, Liang W, Liu C, Lin X, Liu C, Gu Y, Chen W, Liu L, Xu X, Hu Y. A Drosophila single-cell 3D spatiotemporal multi-omics atlas unveils panoramic key regulators of cell-type differentiation. Cell. 2025 Aug 21;188(17):4734-4753.e31. doi: 10.1016/j.cell.2025.05.047. Epub 2025 Jun 26. PMID: 40578340.




